
草地生态系统氮循环关键过程对全球变化及人类活动的响应与机制
2014-01-02 08:12闫钟清齐玉春董云社彭琴孙良杰贾军强曹丛丛郭树芳贺云龙
草业学报 2014年6期
闫钟清,齐玉春,董云社,彭琴,孙良杰,贾军强,曹丛丛,郭树芳,贺云龙
(1.中国科学院地理科学与资源研究所,北京100101;2.中国科学院大学,北京100049)
*全球草地面积约为5000万km2,占全球陆地总面积的33.5%左右,中国草地总面积约为392.8万km2,约占陆地总面积的41%,为现有农田面积的4倍左右[1]。作为陆地生态系统中一个重要的子系统,草地生态系统在全球变化及生态系统功能发挥等方面起着非常关键的作用。氮元素的生物地球化学循环过程作为陆地生态系统最基本的物质循环过程,对全球变化及人类活动存在敏感的响应,并且与碳等其他元素的生物地球化学循环过程密切耦合。SCOPE(国际科联环境问题科学委员会)1998-2001年的科学计划中就着重强调了“氮的循环和转化”,并且IGBP(国际地圈生物圈计划)的许多核心项目也把氮的生物地球化学循环作为其主要研究内容[2]。
在草地生态系统中,氮的生物地球化学途径主要是在大气、土壤、植物、动物、微生物等之间进行,可分为内循环和外循环(图1)。外循环过程是指氮素向生态系统中输入和从生态系统中输出的过程,内循环过程是指氮素化学形态的转变和在系统中不同库间的转移过程[3],其过程主要包括植物对氮素的吸收利用、土壤氮素矿化、硝化和反硝化、铵离子的吸附和释放以及凋落物的分解等。
近年来,由于人口与资源矛盾的加剧,草地生态系统正遭受着越来越严重的人为活动的影响,如过度放牧、草地开垦为农田、火烧等[4]。由于人类活动的强烈影响,导致原本较为封闭的草地氮循环过程逐渐开放,这些干扰行为对草地氮的内外循环过程等均产生了重要的影响。研究氮素循环过程对上述变化的响应过程及机理,不仅对于全面了解草地生态系统氮元素参与的物质循环和能量流动、完善全球变化研究具有重要科学意义,而且也能够满足应对全球气候变化国际行动的国家需求,以及提升我国在全球变化与氮循环领域的理论知识和学术水平、深化氮-碳-气候相互作用研究的科学需求。在草原生态系统中,土壤作为生物量生产最重要的基质,是许多营养的储存库,是动植物分解和循环的场所。因此,本研究重点综述了几种重要的人为活动(放牧、开垦、火烧)及全球变化(全球变暖、氮沉降、大气降水)对土壤中氮素迁移转化动态的影响,探讨了这些因素对草原土壤氮循环的主要影响过程与机制,以便为今后采取合理的草地管理措施,有效防止草原退化、保证草地生态系统的可持续发展提供科学的指导,并对未来的相关研究起到一定的参考作用。
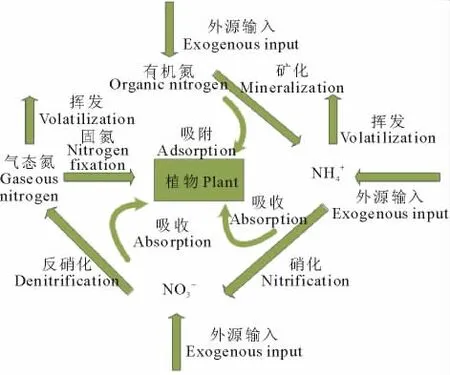
图1 土壤中氮素循环转化简图[5]Fig.1 The soil nitrogen transformation diagram[5]
1 人类活动对草地氮循环关键过程的影响
影响草地生态系统氮循环的人类活动主要包括土地利用方式(放牧、开垦等)和土地利用管理措施(火烧等)的变化等。
1.1 放牧对草地氮循环的影响
放牧是影响草地生态系统氮循环过程的重要人类活动因素[6]。放牧过程中,家畜通过采食、践踏和排泄粪便等影响草地,特别是草地的土壤状况,同时,家畜还通过采食活动加快营养物质的转化、排泄物归还提高土壤营养物质等途径影响草地氮素的循环。因此,放牧对草地土壤氮养分及其循环过程有着显著的影响[7]。与天然状态下的动物采食不同,人类可以通过调节放牧活动的时间、强度、频率等进而对土壤氮循环产生不同的影响效应。
放牧对草地氮素循环的影响主要取决于放牧强度、放牧制度、放牧季节、放牧动物的牧食行为等几个因素,而放牧强度是其中最主要的控制因素[8]。有关放牧强度对土壤氮素储量的影响,大部分研究表明,随着放牧强度的增加,表层土壤有机质和全氮含量明显减少。李春莉[9]在荒漠草原上的研究结果指出,随着放牧强度的增加,草地土壤的氮含量下降。长期的过度放牧已经严重影响了中国北方草地的氮储量[12],虽然进行围栏封育可以显著提高这些草地的氮储量,但是进行至少20年的休牧才可以将一个轻度退化的草地恢复转变为土壤氮储量接近于未受干扰的天然草地[10]。Holland等[11]的研究也得到相同结论,并指出在动物的过度采食下,转运到土壤中的氮下降,同时凋落物的转化率上升。但是也有相关研究表明,并非放牧强度越大土壤氮储量越低。高永恒[13]在对高山草甸生态系统氮素分布格局和循环进行的研究中发现,随放牧强度的增加,0~10cm土壤氮贮量均呈增加的趋势。研究中尽管重度放牧也显著增加了土壤氮贮量,但同时也显著减少了群落盖度,降低了植物地上生物量,这不但不利于畜牧业的持续发展,久而久之还会减少植物向土壤中氮的归还率,带来土壤的退化;同时,随着高寒草原退化程度的加剧,土壤有机质、硝态氮含量、土壤湿度和土壤紧实度也将会减小[14],因此,合理的放牧强度以及频度对于增加或保持土壤氮养分至关重要。
放牧对草地含氮温室气体排放也具有重要影响。Chatskikh等[15]指出草原放牧可能占全球N2O排放量的10%。在放牧过程中,动物排泄物给草原提供大量外加的速效碳、氮底物,从而在一定程度上提高了微生物的活性,促进了N2O的产生[16]。另外,由于牲畜践踏导致放牧草原土壤的压实,在湿润季节使其较围栏草原更容易形成厌氧微区从而增加了厌氧孔隙的比例,有利于反硝化作用产生N2O气体[17]。Flessa等[18]指出在放牧动物尿斑处N2O的释放速率很高,N2O-N损失量相当于排泄物归还氮素的3.2%,估计全球此来源的N2O-N约为1.18Tg/a,因此动物排泄物可能是N2O的重要排放源。然而杜睿等[19]则认为,牲畜的采食与践踏改变了土壤表层结构,从而阻碍了N2O的排放过程,最终放牧行为将会降低草原土壤N2O的排放通量。齐玉春[20]对内蒙古羊草(Leymuschinensis)轮牧草原的研究也发现,与围栏封育相比,放牧并没有显著增加草原土壤N2O的排放。并分析指出,尽管牲畜的排泄物会在一定程度上增加土壤表面的碳氮输入,但羊草轮牧地主要是冬季放牧,土壤温度较低,牲畜排泄物并不会对N2O排放有显著的促进作用,此外,牲畜践踏行为导致的土壤压实,容重提高,孔隙度下降,在增加放牧草原厌氧孔隙的同时,土壤通气性的降低也在一定程度上阻碍了N2O的排放过程。因此,放牧所引起的草原植物、土壤结构以及土壤养分等的变化对土壤N2O通量的影响都具有双重作用,其对最终土壤与大气氮交换通量的影响取决于各种影响的综合对比关系。
Deenik[21]指出,氮矿化潜力是土壤质量的一种量度。不少研究表明,放牧强度越大,氮的矿化速率将会越高。Gao等[22]和Wu等[23]的研究也得出了一致结论,即随着放牧压力的增强总氮矿化率升高,放牧增加了土壤中硝态氮的累积,使氮循环速率加快。分析可能原因是牲畜向土壤中加入了易分解的富含氮素的粪便,此外,放牧通过牲畜的践踏作用还会造成植物残体的破碎,植被凋落物混入土壤有机质、植物盖度下降,土壤表面温度提高等[24],这些环境因素的变化均会对氮的矿化产生刺激作用[25]。而禁牧管理下的植被生长旺盛,土壤水分散失较少,有利于有机化合物络合,为土壤孔隙中的有机物质提供了较多的物理保护,从而限制了土壤微生物等对有机质的矿化分解[26]。相应地,McNaughton等[27]也指出,在草原上停止放牧会导致凋落物氮固持和地上立枯生物量的增加,同时使得土壤中微生物转化率和土壤净氮矿化下降。但也有研究显示,放牧强度若引起草地植被变化则可能会对土壤氮矿化产生相反的效果,Frank等[28]在黄石公园灌丛草地发现,放牧活动与围栏封育相比促进了氮素矿化、提高氮的有效性,同时抑制氮流失。然而,当放牧动物改变了氮对植物的可利用性以及微生物的活性而引起植物群落发生变化时,会导致草地生态系统内的氮素损失量超过输入量,进而抑制氮素的循环。此外,杨小红等[29]分析了内蒙古3个不同放牧强度羊草草原0~30cm表层土壤中矿质氮的浓度特征,结果表明,围封禁牧多年的羊草样地矿质氮含量高于轮牧地和自由放牧地,放牧强度与氮矿化速率负相关。对高寒草甸土壤进行室内培养的结果也显示,在培养初期,禁牧处理下的土壤释放出较多的矿化氮,净氮矿化率随放牧强度的增强而降低[30]。此外,在不同的草地植被类型上进行放牧以及放牧不同的动物品种对草地氮循环过程产生的影响也不一致,在湿润草地的湿生植物群落上进行放牧,净氮矿化率随放牧强度的增加而增大,而在中生植物群落中,净氮矿化率随放牧压力改变无显著变化[31]。
有关放牧活动对硝化过程的影响,Frank等[28]指出在黄石灌丛草原放牧能够多途径地增加土壤微生物的活动、提高硝化酶与反硝化酶活性。李玉中等[32]对羊草草地的研究发现,过度放牧地硝化速率始终高于保护区(无人为扰动)。其主要原因可能是过度放牧致使草地严重退化,牧草生长矮小,进而对土壤中矿质氮的吸收减少,使得土壤中NH4+-N有一定的积累,放牧家畜对所采食植物中的氮只消化吸收一部分,其中60%~90%的养分又以粪尿形式归还草地[33]。当畜尿进入草地后尿素很快又转化为铵,使得土壤中NH4+浓度和pH值有所升高[34],同时,硝化作用产生的NO3--N由于植物利用较少也有一定的积累,因此土壤硝化作用强度较高。但放牧强度对硝化速率的影响存在一个合理范围。高雪峰和韩国栋[35-36]在温带典型草原和荒漠草原进行放牧实验得出,轻度放牧使土壤中氮素转化细菌的数量显著增加,也显著加强了土壤中的硝化作用,促进了土壤中的N素循环。随放牧强度的增加,重度放牧下草原土壤中N素转化细菌数量均显著降低,显著抑制土壤中的固N作用和氨化作用,阻碍了土壤中的N循环进程,但放牧强度对N素转化细菌以及硝化作用的季节性变化规律无明显影响。
从前面的分析可以看出,放牧活动对草地氮循环过程产生的影响会因草地类型和放牧强度的不同而产生较大差异,由于草地土壤生态系统本身的复杂性和弹性,以及不同研究者对放牧强度的界定也不尽相同,因此目前放牧对草地土壤氮循环关键过程的影响并没有明确定论。但可以肯定的是,绝大部分的研究都表明放牧存在一个合理的强度范围,在该范围内可以使草地在提供最大生产力的情况下达到土壤氮养分的保持与可持续利用。
1.2 开垦
草地农垦是除放牧以外,影响草地生态系统的最主要人类活动因素。草地开垦为农田通常会加速土壤中氮的矿化分解速率,导致草地土壤的氮损失和土壤肥力的减弱[37]。
土壤总氮含量受到土地利用方式变化的显著影响[38]。农业开垦使原先草地土壤中被保护着的有机质释放并降解,造成土壤微生物量和有机质含量的不断降低,加速水土流失而导致土壤尤其是表层土壤氮损失。其损失量与气候和土壤类型密切相关。李明峰等[39]对典型温带草原农垦后0~100cm土体不同层次的土壤全氮进行了测定,结果显示,经28年农垦,0~100cm土体中土壤全氮相对于未开垦草原减少约8.8%,平均每年减少0.31%,0~10cm的表层土壤平均减少速率为10~100cm土体的2.9倍。分析原因可能是农垦活动改善了土壤水分条件,土壤硝化作用有所增强,从而降低了土壤中的全氮含量。同时,将小麦(Triticumaestivum)及其秸秆移出农田也大大减少了氮向土壤的回归。此外,草原开垦后的农作措施也会在很大程度上改变土壤结构、理化性质、土壤微生物构成等,从而强烈影响氮库贮量及其与大气的氮交换量。对中国东北地区草地的研究表明,草地开垦为农田后,0~10cm土壤全氮含量显著降低,在10~60cm不同的实验样地表现出不同的规律,而对60 cm土层以下的土壤全氮含量没有影响[40]。对科尔沁退化沙质草地开垦耕种2年后也发现,土壤物理性质和全氮含量均发生显著变化,0~15cm耕作层全氮约下降了28.0%,下降幅度随土层深度而减小。弃耕围封恢复5年后,土壤氮养分的显著提高发生在0~7.5cm土层,7.5cm以下没有变化[41]。相似的,文海燕等[42]对沙质草地的研究也指出在开垦5年后,土壤耕作层已经损失了大量的氮。开垦后有机质的减少使得微生物得不到足够的可利用碳、氮供应,进而降低土壤微生物生物量和活性,氮的矿化和固定也随之受到影响[43]。
草地开垦为耕地一般会带来硝态氮的流失和N2O的释放。一方面是由于开垦后增加了氮肥的施入;另一方面,耕作过程中,人类对土壤的扰动也会显著影响土壤碳、氮的分解矿化[44]。王艳芬[45]指出农垦增强了土壤-植物系统排放N2O的能力。观测期间平均排放通量为2.914μg N/(m2·h),比天然草原高52%,排放通量的季节规律性减弱,整个生长季呈波动性变化,通量变化与天然草原相比趋于平缓。Mosier等[46]发现天然草原进行农耕后的第1年,土壤N2O的排放量约为对照天然草原的8倍,农田弃耕还草后需要8~50年才会恢复到天然草原的水平。刘立新等[47]对草甸草原开垦为农田的研究也表明,土壤N2O通量约比天然草甸草原增加63%。尽管经过20多年的开垦,土壤全氮水平较天然草甸草原已有所降低,但由于农垦过程中人类的农作活动加剧了对土壤的扰动,同时每年1次的基肥施入与小麦秸秆的种前还田仍会在一定程度上增加土壤微生物硝化作用和反硝化作用的反应底物浓度,增加N2O的气态损失。王跃思等[48]指出农垦显著增加了草甸草原N2O的排放通量,农垦麦田生长期内N2O平均排放通量比原始草原高3倍,但农垦并未改变N2O的季节排放变化形式。Machefert等[49]在对欧洲不同土地利用类型N2O排放定量测定结果的总结中指出,相比于森林和草地生态系统,农田生态系统具有最高的N2O排放量,因此将草地生态系统开垦为农田后势必会增加N2O这一温室气体的排放。但是不同的开垦过程也会产生不同的影响,Roth等[50]比较研究了德国北莱茵-威斯特法伦州草地分别利用机械翻耕或者撒入化学除草药剂免耕后撒入玉米(Zeamays)种子转化为农田给它们带来的影响,N2O释放量机械翻耕转化草地大于化学转化未翻耕草地,大约分别为未开垦草地的2.5和2.0倍。2种转换方式都促进硝态氮流失污染地下水并间接促进了N2O的排放,而且机械开垦过程的影响要大于化学开垦。除了转化过程的影响,不同的草地年龄和类型也会对开垦后的硝酸盐淋失产生不同影响,Eriksena等[51]对不同年龄不同植物类型草地进行开垦后研究硝态氮淋失情况发现,1年草地耕作后的硝酸盐淋失量平均为11kg N/hm2,低于2年草地的24 kg N/hm2。相同年龄不同植物类型草地开垦后也有显著差别,耕作后三叶草(Trifolium)和黑麦草(Loliumperenne)草地硝态氮分别损失了10和29kg N/hm2。此外,硝酸盐淋失量加权平均在开垦1年后为8.5mg N/L,第2年则降到了4.5mg N/L,而这些差异在进行适量的氮肥添加后可以进行弥补。
草地开垦为农田会增加温室气体的排放并可能造成地下水的污染,但是退耕还草却往往需要更长的时间,因此,在对草地生态系统的管理利用中一定要首先合理规划,在满足生产生活需要的同时,尽可能地维持生态系统健康并且最大限度地降低土地类型的改变对环境所产生的不利影响。
1.3 火烧
火烧常被认为是草原生态系统维持稳定性与生态健康的重要干扰因子,并日益受到重视,是与草原同生同灭的环境要素,草地的不同利用管理方式中,以开垦和火烧的影响最为显著[52]。在草原形成和演变的地质历史过程中,火一直在调控着草原的结构与动态。火烧可以清除枯落物,追加灰分,使地表颜色变黑,并杀死一些生物活体(包括植物种子和昆虫),同时温度对部分生物体产生有效刺激,空出许多小裸地还有一些衍生现象等[53]。火烧作为一种古老的草地改良措施仍然在世界很多地方相沿成习,成为草原管理的一项重要措施,在草地生态系统中,适时、适当地利用火烧可以促进植被更新,保持草地生态系统的稳定性。
火烧可以通过改变土壤养分状况和土壤温度来直接或间接地影响氮素循环。烧荒措施会使原来固定在植被中的氮素全部释放到大气当中,带来土壤全氮含量的降低。研究表明,高寒山区草地土壤火烧3和4年后土壤全氮分别下降51.66%和42.58%[54]。Pereira等[55]对立陶宛寒温带草原样地春季进行火烧发现,在火烧后的2个月内,由火烧产生的疏水性化合物增加了土壤防水性,而土壤有机氮含量与土壤防水性呈显著负相关,因此,火烧地土壤有机氮含量在火烧后的2个月内相比于未烧地明显降低,随后防水化合物会根据不同颗粒大小以不同的速率浸出,这样的低程度火烧由于对草地影响时间较短而被认为对草地生态系统基本不产生影响。
火烧后,尽管大量的氮素以气态形式损失掉,但土壤中的无机氮却有所增加。Wan等[56]对1955-1999年间发表的87份有关火烧对生态系统的影响报告进行整合分析后指出,火烧显著增加了土壤NH4+-N(94%)、NO3--N(152%)含量,对土壤总氮含量和浓度没有影响。但NH4+-N、NO3--N含量明显受到火烧类型和火烧后的时间不同而表现出不同的时空格局变化。NH4+-N在火烧后增加近2倍,而在1年后便可以恢复至火烧前的水平,NO3--N在火烧后短期内变化较小,约为24%左右,而在半年或1年后达到最高增加3倍左右,之后呈下降趋势。此外,氮素对于火烧的响应也因植被类型等的变化而有所差异。
火烧对N素可利用性的影响程度因火烧强度、频度和火烧时间的不同也有所不同,轻度火烧的样地有机质覆盖度显著高于重度火烧样地,有机质含量、全N、年度N矿化估计值也有相似对比关系。另外,火烧频度高的处理似乎倾向于产生较高的净N矿化速率和固持速率。李玉中等[57]对东北羊草草地所做的火烧实验也表明,降水量相对丰沛情况下,适时及适当频次的火烧可增加返青后一定时期内土壤氮的总矿化与硝化速率;同时,火烧后一些固定在凋落物中的养分以无机态补充到土壤中。Marion等[58]发现,不同的火烧程度会影响灰烬的产生质量和数量,随着火势加强灰烬中所含NH4+-N、NO3--N含量也会出现下降。火烧只是增加了表层土壤(0~5 cm)NH4+-N、NO3--N的可利用性,随火势增强,土壤当中的 NH4+-N增加、NO3--N减少,这或许是火烧后硝化速率变化的影响。Andriesse和Koopmans[59]在美国落基山脉草原群落也发现在草地火烧1年后氮的矿化率有明显升高。在火烧后的第1个生长季土壤矿质氮总量火烧地大于未烧地,火烧1年后用乙炔法测定固氮酶的活性有所降低,并认为这是由无机氮浓度的升高而引起的。有说法认为草原上几年1次的周期性火烧,土壤有机物质被大量消耗,降低了氮素循环的速度。北美草地研究者们指出,在长期火烧制度下,土壤中可矿化氮的数量减少[60],切萨皮克湾的浅沼泽草丛火烧后也发现NH4+-N出现降低[61],火烧还显著降低了内蒙古典型草原土壤的硝化作用[62]。在阿根廷南部草原进行的火烧实验得出,火烧通过增加种内植物组织中的C/N而降低氮循环速率[63]。在堪萨斯州高草草原,测定了1994年长期不火烧与生长季进行火烧的高地和低地发现,火烧和地形均会影响土壤氮素的有效性[64]。氮矿化率在未烧地高于火烧地,而且高地矿化速率比低地高5倍左右。
2 氮素循环对全球变化的响应
目前,全球变化已成为前所未有的一种大尺度的环境干扰压力。由人类活动导致的氮沉降增加、温室气体排放引起的全球气温升高以及降雨频率与格局的变化均使得陆地生态系统的生境处于一种连续性的被干扰状态,从而显著影响生态系统的结构和功能。Wigley和Raper[65]根据IPCC第3次评估报告的预测结果并结合其他相关资料分析认为,在气候变化条件下,1990-2100年间全球平均气温将升高1.7~4.9℃,降水格局也将发生±10%的变化[66]。因此,研究这些变化对生态系统氮素循环的影响及其主要调控机理,对深刻理解草地生态系统氮素动态与全球变化的关系,准确预测未来全球变化情景下草地氮循环过程的可能变化意义重大。
2.1 CO2浓度升高与全球气候变暖
Redfield[67]早在1958年的研究中就发现,在海洋浮游植物和细菌中,碳和养分的比例几乎是一个常数,并且一种生物地球化学元素的变化必然导致其他相关元素的变化。近几十年来,工业化发展造成了CO2浓度的显著增加并引起全球气候变暖,从1880年到1990年间,大气中CO2浓度已由280μmol/mol上升到355μmol/mol,并且每年仍以1.8%的速度继续升高,预计到21世纪中期,大气CO2浓度将达到700μmol/mol左右[68]。IPCC第四次评估报告指出,最近100年(1906-2005年)地表平均温度增加0.74℃(0.56~0.92℃)[69]。在这种背景下,基于碳氮元素变化之间的内在联系,不断升高的CO2浓度及其所带来的全球气候变暖势必会对土壤氮素转化产生直接或间接的影响,CO2浓度升高对氮循环过程的影响已经成为全球变化研究的热点问题之一。
碳和氮之间的相互作用会调节氮的循环过程对不断升高的CO2浓度的响应,从而调控植物对可利用氮素的吸收[70]。比如,Luo等[71]的研究就证明CO2浓度增高会增加不稳定碳的含量,这些不稳定碳通过根部分泌以及根的转化会增加微生物对氮的需求,从而增加植物和土壤微生物之间对可利用氮的竞争,导致生态系统过程的氮限制现象。CO2浓度的升高会影响氮素的可利用性,Niklaus等[72]在瑞士西北部营养贫乏的草原发现NO3--N浓度随CO2浓度增加有所降低。而Müller等[73]在不考虑植物氮吸收的情况下,进行了连续6年15N标记的实验室模拟研究,结果表明,伴随着CO2浓度的不断增加,土壤氮素氨化作用转化的铵盐随之增加,这样一个转化的优势在于铵盐对于氮素流失的反应较小,这可以在CO2浓度不断增加的情况下增加草地生态系统对氮的保留与有效利用。
CO2浓度增加会显著影响土壤N2O的排放。Barnard等[74]通过对欧洲温带草原的研究认为,CO2浓度增加会显著降低土壤硝化酶和反硝化酶活性,进而减少土壤N2O气体排放,其中硝化酶比反硝化酶更容易受到CO2浓度增加的影响。Kammann等[75]却得出了相反的结论,通过开放式CO2浓度升高实验(free air carbon dioxide enrichment,FACE)对一个氮限制温带草原进行连续9年的N2O值测定,结果表明,在实验之前不同样地之间N2O排放是没有差异的,从1998-2006年,N2O释放量在高CO2浓度比低CO2浓度情况下增加了2倍多,分别为0.90和2.07kg N2O-N/(hm2·a)。在CO2浓度不断增加下,若固碳作用得不到相应提高,温带草原可通过对N2O排放的正反馈转化为温室气体源。Baggs等[76]对瑞士黑麦草草地利用FACE试验研究了N2O的产气途径,结果发现,在外界CO2浓度超过600μL/L时,反硝化过程占主导地位;而CO2浓度为360μL/L时,硝化过程占主导地位,增加地下部分的碳分配会促进土壤反硝化活性,进而增加N2O通量。而且值得一提的是,随着CO2浓度增高,N2/N2O也随之升高,由此说明之前只是测定N2O产生量会大大低估反硝化作用的氮素损失。
此外,由CO2浓度增加而引起的温度升高还会影响参与氮循环过程的功能微生物特性,进而改变由此驱动的氮循环过程。Ineson等[77]模拟研究了全球气候变暖对土壤氮动态的影响,得出升温不仅增加了氮矿化,并且增加了矿质氮的植物吸收。Barrett和Burke[78]沿温度梯度研究了美国半干旱草原土壤有机质含量对N的固定潜力与转化的影响时发现,N的固定潜力与转化能力随有机质含量的增加而增加,而土壤C/N与年均温呈负相关,因此可以得到氮的迁移转化速率会随着温度的升高而降低。然而温度因子并不是单独对氮循环产生影响,Loiseau和Soussana[79]指出,若只提高CO2浓度会使土壤大团聚体中标记氮的固定增加,草地对氮的吸收减少。相比之下,CO2浓度增加并且温度提高3℃条件下,却促进了氮的可利用性,提高了草地对氮的吸收。可见温度与CO2浓度变化之间在影响氮素循环方面存在一定的耦合作用关系。
2.2 降水变化
水分是影响土壤氮素可利用性的重要环境因子。水分既是氮素溶解的媒介,又是氮素转移的载体,氮素的循环和利用情况与降水因子密切相关。利用IPCC第四次评估报告提供的13个气候系统模式对中国未来100年气候变化的预测结果表明:21世纪中国气候预估显著变暖、变湿,其中70%以上模式的预估年降水量增加范围为14~155mm,且降水的增加主要集中在我国北方草地生态系统的主要分布区域[80]。因此,在全球及我国降水格局发生变化的背景下,草地土壤氮素的迁移转化过程势必将会受到很大的影响。
降水格局的改变对草地生态系统氮损失有着重要影响。全球温室气体浓度的增加会潜在的扩大降水分布的强度和季节可变性,在对北美中纬度地区的稀树草原长达6年的温度降水变化模拟试验中发现,土壤总氮含量维持在59~76g N/m2之间,而土壤水分含量较低的干旱季节土壤氮含量较低,这可能是由于水分限制了初级生产力使得氮储存较少[81]。但是若土壤水分含量超过了植物生长需求也会造成氮素的淋失。硝态氮的淋失早在1905年就曾被英国科学家 Warrington[82]注意。Walvoord等[83]发现,在干旱区的土壤亚表层存在着较大的氮库,这表明氮淋溶可能成为生态系统氮损失的潜在过程,该过程主要受降水变化的影响。张兴昌和邵明安[84]提出土壤氮素与降雨的相互作用表现为2种形式,其一,表层土壤氮素在雨滴作用下,向雨水中释放或被雨滴溅蚀;其二,表层土壤氮素特别是硝态氮随雨水在土壤入渗,当降水强度小于土壤入渗率时,表土氮素特别是硝态氮在土壤深层沉积。
降水量变化带来的土壤水分含量的变化主要通过对土壤的通气状况和氧分压的改变而对土壤N2O的产生和排放过程产生重要影响。Schaufler等[85]通过实验室培育测定欧洲草地生态系统N2O排放与湿度的关系时发现N2O与土壤含水量呈正相关。Horváth等[86]对匈牙利半干旱沙地和黄土草地进行灌溉试验发现,在灌溉区和对照区测得的 N2O通量分别为1.56和0.92kg N/(hm2·a)。Abbasi和Adams[87]对放牧草原分别施用了硝态氮肥和铵态氮肥后发现,无论施加铵态氮肥还是硝态氮肥,N2O的通量以及氮的流失量和流失速率都随土壤充水孔隙度(water filled pore space,WFPS)升高而增加。但也有研究表明,硝化速率存在1个临界含水量,在较低的土壤含水量范围内,土壤中硝化速率与土壤含水量呈正相关;含水率在30%~50%范围内时,硝化速率达到最大;超过临界含水量,硝化速率与含水率呈负相关[88]。Norton等[89]对美国怀俄明州2类灌草草原的研究也表明,短期内土壤的干湿变化对N2O排放量的变化具有十分显著的影响,但不同的变化方向,两者的关系存在差异。土壤由干变湿过程中,N2O通量随含水量的增加而上升,从干到水分饱和这个过程中,N2O通量的变化较平稳,呈直线逐渐上升的趋势,而土壤由湿变干过程中最大N2O通量则出现在 WFPS为70%时,而后随着土壤WFPS含量的减少,N2O通量急速下降。土壤含水量一方面除了通过影响矿质氮的迁移,进而影响N2O的产生量及其产生途径外,另一方面,由于N2O在空气中的扩散速率约为水中的104倍[90],因此,土壤水分还可以通过影响土壤的通气状况而对土壤中N2O从产生部位向大气中的扩散与传输产生重要影响。
在干旱半干旱草原中,降水变化也是影响土壤氮素矿化速率、NH4+和NO3-浓度的分布及其对微生物有效性的重要驱动因子。在干旱季节,表土矿质氮的含量保持恒定,而在雨季开始后,降水的格局决定了氮的可获得性。雨量较小时,即导致矿化发生,约5d后,表土层的矿质氮含量将加倍;相反,在雨量较大时,表土矿质氮的含量在开始2个星期就减小了50%~70%,原因可能是大量降水增加地表径流引起矿质氮流失。另外一个原因还可能是矿质碳库的减小引起土壤中碳源缺乏,导致参与矿化的土壤微生物无法获取能量,而氮的矿化被认为是通过微生物而与碳的流动相连接的[91]。Calderon等[92]通过耕作和未耕作土壤中测定氮素矿化作用实验发现,由于微生物对翻耕所产生的土壤湿度变化的响应,土壤含水量的减少使得氮素的矿化降低了15%。Xiang等[93]对加利福尼亚草原进行干湿交替,发现经过多个周期之后,表层矿化速率相应地也增加了1.5倍,亚表层氮矿化率增加了大约8倍,与不断进行湿润处理的增加量相当。
2.3 氮沉降
大气氮沉降的研究始于英国洛桑试验站[94],它是氮素生物地球化学循环中的重要环节。化石燃料燃烧、氮肥施用、畜禽养殖等人为活动,致使过去的1个世纪内大气中的活性氮化合物增加了约3倍[95]。近年来,我国大气氮沉降也在逐年上升,从1980年至2010年,中国总的氮沉降量平均以每年0.41kg N/hm2的速率持续增加[96]。氮沉降带来生态系统氮输入增加以及土壤环境的显著变化,并与气候变化一起,进一步导致和加剧氮循环过程的改变。此外,大气氮素干湿沉降等也是自然生态系统一项重要的养分资源,通过综合管理,充分利用这一环境养分资源是实现自然生态系统可持续发展的关键[97]。
作为生态系统氮输入的重要来源之一,大气氮沉降的变化将直接影响土壤的氮素储量及有效态氮含量。在中国温带典型草原发现,随着氮沉降速率增加,土壤的总氮含量,NH4+-N和NO3--N含量均有所增加[98]。Zhang等[99]在中国东北部松嫩草甸草原添加10g/(m2·a)的硝酸铵模拟氮沉降的试验中发现氮添加增加了土壤总氮含量和有效氮含量。但Verena等[100]在阿尔卑斯山海拔2000m左右的亚高山草原做的模拟氮沉降实验却发现50kg N/(hm2·a)氮添加并没有影响土壤总氮含量,这可能跟草原存在氮限制状态有关。在英国的酸性草原和石灰石草原所做的氮添加实验发现140kg N/(hm2·a)的年氮沉降速率对土壤总氮含量也并无影响[101]。在欧洲酸性草原通过添加梯度范围在2~44kg N/(hm2·a)之间的模拟氮沉降发现草地C/N增加,这说明相比于之前预期的氮会随着碳的增加而增加,更确切的是可以得到相对于氮素的增加,碳的相应含量会增加更多。但是土壤中可提取的硝酸盐和铵的浓度并没有受到氮沉降的影响,同时指出造成这一现象的机制还有待于更多的研究[102]。
陆地生态系统活性氮含量的剧增可能会增强硝化作用和反硝化作用,进而加剧土壤N2O气体的排放。有研究者对109个已发表的实验结果进行了统计,发现生态系统中由于氮的沉降造成N2O排放增加216%[103]。许多试验都证实氮添加后N2O排放量的变化取决于土壤中的NO3-和NH4+的有效性及土壤理化性质等因素[104],李英臣等[105]在实验室培养条件下研究净氮输入分别为1,2和5mg/g水平时对草甸沼泽土反硝化损失和N2O排放的影响,结果显示,培养期间3种氮输入水平的N2O排放速率平均值分别为12.55,7.59和4.04 μg/(kg·h),明显高于对照的0.09μg/(kg·h),但高氮输入会对N2O排放速率产生一定的抑制作用,且随着梯度增大而显著加强。对波多黎各西部撂荒25年草原的研究表明,氮添加对3种草地土壤N2O的排放均有不同程度增强,有0.8%~3.3%的氮肥以N2O的形式进入了大气[106],然而也有氮添加后对草地土壤硝化与反硝化过程无明显影响的例子。例如,Stephan和Karl[107]在德国南部的草地生态系统进行的为期2年的模拟氮沉降实验(200kg含氮牛粪浆/hm2·a)结果表明,氮添加对N2O的排放不产生显著影响,N2O释放在施肥地区和不施肥地区均较小,并指出快速氮矿化与表土密集根部的吸收阻碍了氮损失。生态系统是否处于氮限制对于氮沉降增加后N2O的排放变化也起到了重要影响,Kim等[108]选择2个具有不同本底土壤氮含量(高氮地总氮含量约为低氮地的2倍)但土壤物理性质没有差别的样地来比较两者对氮沉降的响应差异,结果表明,2种样地氮添加均增加了NH4+-N和NO3--N含量,并刺激了N2O气体的释放,但高氮背景样地N2O排放显著高于低氮背景样地,气体排放量分别占施氮总量的1.66%和0.69%,并得出N2O和NO3--N浓度之间存在正相关关系(R2=0.80,P<0.0001)。
氮素添加后的NH3挥发也是氮损失的一个重要过程,并且不同的添加类型对NH3挥发的影响存在一定差异。Catchpooir等[109]指出,在温度相对较高的环境条件下,将尿素施到放牧场上,NH3的挥发损失量最高可达42%。Vander Weerden和Jarvis[110]在美国北威客的黑麦草草地进行的氮添加实验表明,草地添加尿素后氨挥发损失为添加量的12%~46%。而施加硝酸铵和硝酸钙则具有较小的氨挥发量,不到添加量的1%。对科尔沁东部沙质草地氮添加短期试验结果表明,氮添加处理的氨挥发累积量显著高于对照处理,且氨挥发速率表现出先升高后降低的趋势,直至与对照样地无显著差异[111]。
很多研究指出长期的氮沉降能促进土壤有机氮的矿化速率。杨江龙[112]研究表明,施氮草地土壤中氮素的矿化作用大都高于未施氮的同一类型草地,氮素添加促进原来土壤有机氮分解、释放。在中国东北部的松嫩草甸草原用添加10g/(m2·a)的硝酸铵模拟氮沉降,也发现氮添加增加了土壤氮素矿化速率[113]。白洁冰等[114]研究氮素输入对青藏高原3种高寒草地土壤氮矿化的影响发现,氮素输入显著促进了高寒草原土壤氮矿化速率,土壤氮矿化速率随着氮处理剂量的增加而升高,对于氮素添加促进土壤的氮矿化作用这一现象,推测可能是由于施入的无机氮被微生物固定,从而促使原来有机氮的矿化和释放。但氮素添加达到一定程度后也会限制矿化作用,张璐等[115]对内蒙古典型草原羊草群落开展了长期的氮素添加试验,也得出氮素添加显著改变了土壤的累积氮矿化量,但高氮处理下的累积氮矿化量较低氮处理低,净矿化作用在中等氮沉降水平达到最高。氮矿化作用不仅受到氮素添加量的影响,还与土壤氮素本底值密切联系。Gundersen等[116]的野外试验研究发现,只有在氮限制地区氮输入增加才会提高净矿化速率,而在氮循环速率较快的地区,进一步施氮反而会减慢土壤氮的净矿化作用。除此之外,由于模拟氮沉降实验一般都是一段时间内进行氮素添加,不会永久持续进行,因此,氮素添加对土壤氮素矿化的作用存在一个时间范围。Haynes[117]通过3年草原施肥实验发现,土壤有机氮的净矿化速率会随着肥料氮的添加而上升,而这种增加持续时间较短。Aber等[118]指出添加的氮在短期内与有机物质结合会降低土壤C/N,从而加速了土壤有机物的分解和养分的释放过程,但在长期持续施氮条件下,土壤氮素的总矿化作用虽有增加,但其净矿化速率从先期的峰值开始出现下降,逐渐接近或低于对照值。
氮沉降增加对草地土壤氮素循环过程的影响与草地本身的氮素基础水平、降水条件等均有着密切的关系。不同氮输入量与氮输入类型对土壤氮的矿化、淋溶以及气态损失也存在直接的影响,此外,不同类型的草地群落对氮的需求量不同,也会间接影响土壤氮的迁移、转化、分布和利用。因此,今后应加大不同草地类型、不同氮输入量及氮输入类型下影响效应的比较研究,以便更准确全面、科学的评价氮沉降增加可能造成的不良环境影响,为氮沉降增加背景下草地生态系统应对措施的制定提供理论依据。
3 展望
从对现有文献结果的分析中可以看出,以往氮循环相关研究多集中在氮肥施用较多的农田生态系统,对草地以及森林等自然生态系统研究较少。同时,对草地生态系统而言,由于氮循环过程及其影响因素的复杂性,目前的研究一般通常只局限于氮循环的某一个环节,基于整个循环过程以及探讨多个全球变化因子耦合对氮循环过程影响的定量研究则很少涉及,且目前我国仍缺少长期、系统、大规模的相关科学研究计划,对相关机制的分析也多局限于环境因子与氮循环过程变化的数理统计分析,缺乏对其微生物学驱动机制的深入探讨,相关研究亟待深入与完善。今后的研究工作应重点在以下几个方面有所加强。
1)建立标准化的研究体系,按照相关标准系统监测草地生态系统氮循环对人类活动及全球变化的响应。通过过去长期的科学实践,多尺度的长期野外观测、人工控制实验、生态系统过程模拟、模型模拟等技术途径的有机融合已被证明是全球变化情景下区域尺度元素生物地球化学循环过程研究的有效手段。本文所综述的实验绝大多数为独立的野外实验研究,因本底差异、实验设计、自然条件限制等诸多方面的因素,使得研究结果差异性大,规律性不明显,不同研究结果的可比性差。由于土壤氮素循环过程是一个相互联系、相互作用的多途径的统一体,需要采用综合的系统分析方法进行研究,因此今后应当借鉴类似包括FLUXNET在内的全球尺度监测体系,通过协商探讨设定相关的实验标准,形成更为规范系统的全球变化研究技术方法体系范式,以最大限度地减少因实验设计本身差异所导致的结果偏差,同时也有利于研究数据的共享和管理。
2)开展草地生态系统的长期监测研究。与农田生态系统拥有大量长期人工控制研究有所不同的是,草地生态系统长期控制实验仍相当缺乏。单靠短期试验是无法深刻把握其变化规律的,有时甚至会得到相反的结论,只有建立起长期的野外试验研究平台,探讨长时间周期内各种复杂外因对研究对象的影响,才能对变化本身做出更为科学的评价。
3)加强不同草地类型的对比研究。我国草地类型也多种多样,共包括18大类、38亚类,上千种类型。由于不同的草地类型所处气候带,物种构成和土壤基质(如土壤动物和土壤微生物种类、土壤的结构、pH值以及土壤的有机质含量等)均存在一定不同,因此,其氮循环过程对人类活动与环境变化的响应也势必存在较大差异,只有广泛开展不同草地类型的对比研究才能使我们对草地的相关认知更加全面和系统。
4)加强相关微生物生态学机制的深入探讨。微生物是土壤碳氮转化的主要驱动者,是对环境变化最为敏感的生命指标,在土壤碳氮循环及其对自然和人为干扰响应中具有十分重要的表征功能。人类活动与全球变化会直接或间接地影响土壤微生物活性,特别是微生物介导的生物地球化学循环过程。但目前相关研究仍极为薄弱。土壤微生物数量,群落结构和多样性等是如何与气候及人类扰动相互关联,又是如何驱动土壤氮转化,进而影响生态系统氮过程的仍有待进一步深入探索。
5)完善相关过程机理模型。我国的氮循环模型研究起步比欧美等发达国家晚,虽然近些年发展非常迅速,先后引进和改良了多个陆地生态系统氮循环模型,但目前国内对生态系统氮循环模型的研究主要集中在农田,对草地生态系统氮循环的研发还处于起步阶段,还没有建立完整的、适宜于草地的生理生态学参数体系,对于全球不同变化方式下的氮循环过程改变以及对于生态系统与流域尺度上氮循环的模拟能力还有待于进一步完善。
6)加强全球变化多因子以及环境变化与人类活动的耦合影响效应研究。人类活动与全球变化共同影响着氮循环过程,并且全球变化多因子之间也是相互影响相互制约的。若单纯探讨某一因子变化对生态系统氮循环过程影响,其研究结果在另一个因子背景发生变化后就很难反映其影响的真实状况,这就在一定程度上限制了研究成果的应用,同时也很难充分发挥不同因子之间的协同作用。因此,应当形成系统的研究网络体系,对包括气候变暖、降水变化、氮沉降等在内的全球变化因素进行大尺度交互模拟研究,以便更准确地预测和有效调节生态系统的氮循环过程。
[1]李博.我国草地资源现状,问题及对策[J].中国科学院院刊,1997,1:49-51.
[2]Gruber N,Galloway J N.An earth-system perspective of the global nitrogen cycle[J].Nature,2008,451:293-296.
[3]陈佐忠,汪诗平.中国典型草原生态系统[M].北京:科学出版社,2000:228-246.
[4]彭琴,董云社,齐玉春.氮输入对陆地生态系统碳循环关键过程的影响[J].地球科学进展,2008,23(8):874-882.
[5]周志华,肖化云,刘丛强.土壤氮素生物地球化学循环的研究现状与进展[J].地球与环境,2004,32(3-4):21-26.
[6]刘天增,南志标.放牧对草地的作用[J].草业科学,2011,28(6):951-958.
[7]张成霞,南志标.放牧对草地土壤理化特性影响的研究进展[J].草业学报,2010,19(4):204-211.
[8]王忠武.载畜率对短花针茅荒漠草原生态系统稳定性的影响[D].呼和浩特:内蒙古农业大学,2009.
[9]李春莉.放牧对短花针茅草原及糙羊茅草原植被和土壤影响的研究[D].呼和浩特:内蒙古农业大学,2008.
[10]He N P,Yu Q,Wu L,etal.Carbon and nitrogen store and storage potential as affected by land-use in aLeymuschinensisgrassland of northern China[J].Soil Biology and Biochemistry,2008,40(12):2952-2959.
[11]Holland E A,Parton W J,Detling J K,etal.Physiological responses of plant populations to herbivory and their consequences for ecosystem nutrient flow[J].American Naturalist,1992,140:685-706.
[12]王利民,姜怀志,姚纪元,等.我国北方草地的现状和可持续发展对策[J].家畜生态,2004,25(2):4-7.
[13]高永恒.不同放牧强度下高山草甸生态系统碳氮分布格局和循环过程研究[D].成都:中国科学院成都生物研究所,2007.
[14]周华坤,赵新全,温军,等.黄河源区高寒草原的植被退化与土壤退化特征[J].草业学报,2012,21(5):1-11.
[15]Chatskikh D,Olesen J E,Berntsen J,etal.Simulation of effects of soils,climate and management on N2O emission from grasslands[J].Biogeochemistry,2005,76:395-419.
[16]Yamulki S,Jarvis S C,Oven P.Nitrous oxide emission from excreta applied in a simulated grazing pattern[J].Soil Biology and Biochemistry,1998,30:491-500.
[17]旭日.内蒙古草原锡林河流域N2O气体交换过程模拟与尺度转换[D].北京:中国科学院大气物理研究所,2002.
[18]Flessa H,Dörsch P,Beese F,etal.Influence of cattle wastes on nitrous oxide and methane fluxes in pasture land[J].Journal of Environment Quality,1996,25:1366-1370.
[19]杜睿,王庚辰,吕达仁.放牧对草原土壤N2O产生及微生物的影响[J].环境科学,2001,22(4):11-15.
[20]齐玉春.内蒙古温带草地生态系统生物地球化学循环中主要温室气体通量与碳平衡[D].北京:中国科学院研究生院,2003.
[21]Deenik J.Nitrogen mineralization potential in important agricultural soils of Hawaii[J].Soil Crop Management,2006,15:1-5.
[22]Gao Y H,Luo P,Wu N,etal.Impacts of grazing intensity on nitrogen pools and nitrogen cycle in an alpine meadow on the eastern Tibetan plateau[J].Applied Ecology & Environmental Research,2008,6(3):69-79.
[23]Wu H H,Dannenmann M,Fanselow N,etal.Feedback of grazing on gross rates of N mineralization and inorganic N partitioning in steppe soils of Inner Mongolia[J].Plant & Soil,2011,340(1-2):127-139.
[24]Han G D,Hao X Y,Zhao M L,etal.Effect of grazing intensity on carbon and nitrogen in soil and vegetation in a meadow steppe in Inner Mongolia[J].Agriculture Ecosystem & Environment,2008,125:21-32.
[25]Rossignol N,Bonis A,Bouzill J B.Consequence of grazing pattern and vegetation structure on the spatial variations of net N mineralization in a wet grassland[J].Applied Soil Ecology,2006,31:62-70.
[26]Peter S,Erik S J.Mineralization-immobilization and plant uptake of nitrogen as influenced by the spatial distribution of cattle slurry in soils of different texture[J].Plant and Soil,1995,173(2):283-291.
[27]McNaughton S J,Ruess R G,Seagle S W.Large mammals and process dynamics in African ecosystems[J].BioScience,1988,38:794-800.
[28]Frank D A,Groffman P M,Evans R D,etal.Ungulate stimulation of nitrogen cycling and retention in Yellowstone Park grasslands[J].Oecologia,2000,123(1):116-121.
[29]杨小红,董云社,齐玉春,等.锡林河流域羊草草原暗栗钙土矿质氮动态变化[J].地理研究,2005,3:387-393.
[30]陈懂懂,孙大帅,张世虎,等.青藏高原东缘高寒草甸土壤氮矿化初探[J].草地学报,2011,19(3):420-424.
[31]Xu Y Q,Li L H,Wang Q B,etal.The pattern between nitrogen mineralization and grazing intensities in an Inner Mongolian typical steppe[J].Plant and Soil,2007,300(1-2):289-300.
[32]李玉中,祝廷成,Redmann R E.三种利用类型羊草草地氮总矿化,硝化和无机氮消耗速率的比较研究[J].生态学报,2002,22(5):668-673.
[33]Haynes R J,Williams P H.Nutrient cycling and soil fertility in the grazed pasture ecosystem[J].Advances in Agronomy,1993,49:119-199.
[34]Luo J,Tillman R W,Ball P R.Grazing effects on denitrification in a soil under pasture during two contrasting seasons[J].Soil Biology and Biochemistry,1999,31:903-912.
[35]高雪峰,韩国栋.放牧对羊草草原土壤氮素循环的影响[J].土壤,2011,43(2):161-166.
[36]高雪峰,韩国栋.利用强度对荒漠草原土壤氮循环系统的影响[J].干旱区资源与环境,2011,25(11):165-168.
[37]李凌浩,陈佐忠.草地群落的土壤呼吸[J].生态学杂志,1998,17(4):45-51.
[38]Liu X R,Dong Y S.Drivers of soil net nitrogen mineralization in the temperate grasslands in Inner Mongolia,China[J].Nutrient Cycling in Agroecosystems,2010,87:59-69.
[39]李明峰,董云社,齐玉春,等.温带草原土地利用变化对土壤碳氮含量的影响[J].中国草原,2005,27(1):1-6.
[40]Ding F,Hu Y L,Li L J,etal.Changes in soil organic carbon and total nitrogen stocks after conversion of meadow to cropland in Northeast China[J].Plant and Soil,2013,373(1-2):659-672.
[41]苏永中,赵哈林,文海燕.退化沙质草地开垦和封育对土壤理化性状的影响[J].水土保持学报,2002,16(4):5-8,126.
[42]文海燕,赵哈林,傅华.开垦和封育年限对退化沙质草地土壤性状的影响[J].草业学报,2005,14(1):31-37.
[43]仲延海,包青海.锡林河流域合理割草制度的研究[J].中国草地,1999,3:28-41.
[44]耿远波,章申,董云社,等.草原土壤的碳氮含量及其与温室气体通量的相关性[J].地理学报,2001,12(6):938-942.
[45]王艳芬.人类活动干扰下草原温室气体地-气交换特征及碳平衡研究[D].北京:中国科学院植物研究所,2001.
[46]Mosier A R,Parton W J,Valentine D W,etal.CH4and N2O fluxes in the Colorado short grass Steppe 2long-term impact of land use change[J].Global Biogeochemical Cycles,1997,11(1):29-42.
[47]刘立新,董云社,齐玉春.锡林河流域生长季节不同草地类型根系呼吸特征研究[J].环境科学,2003,27(12):2376-2381.
[48]王跃思,纪宝明,黄耀,等.农垦与放牧对内蒙古草原N2O、CO2排放和CH4吸收的影响[J].环境科学,2001,22(6):7-13.
[49]Machefert S E,Dise N B,Goulding K W T,etal.Nitrous oxide emission from a range of land uses across Europe[J].Hydrology and Earth System Sciences,2002,6:325-338.
[50]Roth G,Flessa H,Helfrich M,etal.Impact of two different types of grassland-to-cropland-conversion on dynamics of soil organic matter mineralization and N2O emission[J].Geophysical Research Abstracts,2012,14:22-27.
[51]Eriksena J,Pedersenb L,Jrgensenc J R.Nitrate leaching and bread-making quality of spring wheat following cultivation of different grasslands[J].Agriculture,Ecosystems & Environment,2006,116(3-4):165-175.
[52]李愈哲,樊江文,张良侠.不同土地利用方式对典型温性草原群落物种组成和多样性以及生产力的影响[J].草业学报,2013,22(1):1-9.
[53]周道玮.火烧对草地的生态影响[J].中国草地,1992,2:74-77.
[54]周瑞莲,张普金,徐长林.高寒山区火烧土壤对其养分含量和酶活性的影响及灰色关联分析[J].土壤学报,1997,34(1):89-96.
[55]Pereira P,Ùbeda X,Mataix-Solera J,etal.Short-term spatio-temporal spring grassland fire effects on soil colour,organic matter and water repellency in Lithuania[J].Solid Earth Discussions,2013,5(2):2119-2154.
[56]Wan S Q,Hui D F,Luo Y Q.Fire effects on ecosystem nitrogen pools and dynamics:A meta-analysis[J].Ecological Application,2001,11:1349-1365.
[57]李玉中,祝廷成,李建东,等.火烧对草地土壤氮总矿化,硝化及无机氮消耗速率的影响[J].应用生态学报,2003,14(2):223-226.
[58]Marion G M,Moreno J M,Oechel W C.Fire severity,ash deposition,and clipping effects on soil nutrients in shaparral[J].Soil Science Society of America Journal,1991,55(1):235-240.
[59]Andriesse J P,Koopmans T T.Fire effects on nitrogen mineralization and fixation in mountain shrub and grassland communities[J].Journal of Range Management,1984,37(5):402-405.
[60]Blair J M.Fire,N availability,and plant response in grasslands:a test of the transient maxima hypothesis[J].Ecology,1997,78:2359-2368.
[61]Matamala R,Drake B G.The influence of atmospheric CO2enrichment on plant-soil nitrogen interactions in a wetland plant community on the Chesapeake Bay[J].Plant and Soil,1999,210:93-101.
[62]Zhou L S,Huang J H,Lu F M,etal.Effects of prescribed burning and seasonal interannual climate variation on nitrogen mineralization in a typical steppe inner Mongolia[J].Soil Biology and Biochemistry,2009,41:796-803.
[63]Harris W N,Morettob A S.Fire and grazing in grasslands of the Argentine Caldenal:Effects on plant and soil carbon and nitrogen[J].Acta Oecologica,2007,32:207-214.
[64]Turner C L,Blair J M,Schartz R J,etal.Soil N and plant responses to fire,topography,and supplemental N in tallgrass prairie[J].Ecology,1997,78:1832-1843.
[65]Wigley T M L,Raper S C B.Interpretation of high projections for global-mean warming[J].Science,2001,293:451-454.
[66]Wallace J S.Increasing agricultural water use efficiency to meet future food production agriculture[J].Ecosystems and Environment,2000,82:105-119.
[67]Redfield A C.The biological control of chemical factors in the environment[J].American Scientist,1958,46:205-221.
[68]Houghton R A,Skole D L.Carbon In:The Earth as Transformed by Human Action[M].Cambridge:Cambridge University Press,1990:393-408.
[69]IPCC.Climate Change 2007:Synthesis Report[R].Oslo:Intergovernmental Panel on Climate Change,2007.
[70]Peter B R,Bruce A H,Luo Y Q.Carbon-nitrogen interactions in terrestrial ecosystems in response to rising atmospheric carbon dioxide[J].Annual Review of Ecology,Evolution,and Systematic,2006,37:611-636.
[71]Luo Y,Currie W S,Dukes J S,etal.Progressive nitrogen limitation of ecosystem responses to rising atmospheric carbon dioxide[J].BioScience,2004,54:731-739.
[72]Niklaus P A,Kandeler E,Leadley P W,etal.A link between plant diversity,elevated CO2and soil nitrate[J].Oecologia,2001,127:540-548.
[73]Müller C,Rüttingc T,Abbasi M K,etal.Effect of elevated CO2on soil N dynamics in a temperate grassland soil[J].Soil Biology and Biochemistry,2009,41(9):1996-2001.
[74]Barnard R,Barthes L,Le Roux X,etal.Dynamics of nitrifying activities,denitrifying activities and nitrogen in grassland mesocosms as altered by elevated CO2[J].New Phytologist,2004,162(2):365-376.
[75]Kammann C,Müller C,Grünhage L,etal.Elevated CO2stimulates N2O emissions in permanent grassland[J].Soil Biology and Biochemistry,2008,40(9):2194-2205.
[76]Baggs E M,Richter M,Cadisch G,etal.Denitrification in grass swards is increased under elevated atmospheric CO2[J].Soil Biology and Biochemistry,2003,35(5):729-732.
[77]Ineson P,Benham D G,Poskitt J,etal.Effects of climate change on nitrogen dynamics in upland soils:A soil warming study[J].Globle Change Biology,1998,4:153-161.
[78]Barrett J E,Burke I C.Potential nitrogen immobilization in grassland soils across a soil organic matter gradient[J].Soil Biology and Biochemistry,2000,32:1707-1716.
[79]Loiseau P,Soussana J F.Effects of elevated CO2,temperature and N fertilization on nitrogen fluxes in a temperate grassland ecosystem[J].Global Change Biology,2000,6:953-965.
[80]江志红,张霞,王冀.IPCC-AR4模式对这个21世纪气候变化的情景预估[J].地理研究,2008,27(4):787-799.
[81]Rosenstein R W,Boutton T W,Tjoelker M,etal.Soil C and N Pools in Oak Savanna:Responses to Temperature and Rainfall Manipulation[C].American Geophysical Union,Fall Meeting,2011.
[82]Warrington R.Lost fertility[A].The Production and Loss of Nitrate in the Soil[M].Transaction of the High land and Agricultural Society of Scotland,1905:1-35.
[83]Walvoord M A,Phillips F M,Stonestrom D A,etal.A reservoir of nitrate beneath desert soils[J].Science,2003,302:1021-1024.
[84]张兴昌,邵明安.坡地土壤氮素与降雨、径流的相互作用机理及模型[J].地理科学进展,2000,19(2):128-135.
[85]Schaufler G,Kitzler B,Schindlbacher A,etal.Greenhouse gas emissions from European soils under different land use:effects of soil moisture and temperature[J].European Journal of Soil Science,2010,61(5):683-696.
[86]Horváth L,Grosz B,Machon A,etal.Estimation of nitrous oxide emission from Hungarian semi-arid sandy and loess grasslands;effect of soil parameters,grazing,irrigation and use of fertilizer[J].Agriculture,Ecosystems and Environment,2010,139:255-263.
[87]Abbasi M K,Adams W A.Gaseous N emission during simultaneous nitrification-denitrification associated with mineral N fertilization to a grassland soil under field conditions[J].Soil Biology & Biochemistry,2000,32:1251-1259.
[88]刘巧辉.应用BaPs系统研究旱地土壤硝化-反硝化过程和呼吸作用[D].南京:南京农业大学,2005.
[89]Norton U,Mosier A R,Morgan J A,etal.Moisture pulses,trace gas emissions and soil C and N in cheatgrass and native grass-dominated sagebrush-steppe in Wyoming,USA[J].Soil Biology & Biochemistry,2008,40:1421-1431.
[90]梁东丽,同延安,Ove Emteryd,等.灌溉和降水对旱地土壤N2O气态损失的影响[J].植物营养与肥料学报,2002,8(3):298-302.
[91]Saetre P.Decomposition,microbial community structure,and earthworm effects along a birch spruce soil gradient[J].Ecology,1998,79(3):834-846.
[92]Calderon J F,Louise E J,Scow K M,etal.Microbial response to simulated tillage in cultivated and uncultivated soils[J].Soil Biology and Biochemistry,2000,32:1547-1559.
[93]Xiang S R,Doylec A,Holden P A,etal.Drying and rewetting effects on C and N mineralization and microbial activity in surface and subsurface California grassland soils[J].Soil Biology and Biochemistry,2008,40(9):2281-2289.
[94]Goulding K W T,Bailey N J,Bradbury N J,etal.Nitrogen deposition and its contribution to nitrogen cycling and associated soil processes[J].New Phytologist,1998,139(1):49-58.
[95]Galloway J N,Townsend A R,Erisman J W,etal.Transformation of the nitrogen cycle:recent trends,questions,and potential solutions[J].Science,2008,320:889-892.
[96]Liu X J,Zhang Y,Han W X,etal.Enhanced nitrogen deposition over China[J].Nature,2013,494:459-462.
[97]刘学军,张福锁.环境养分及其在生态系统养分资源管理中的作用:以大气氮沉降为例[J].干旱区研究,2009,26(3):306-311.
[98]Zhang X M,Han X G.Nitrogen deposition alters soil chemical properties and bacterial communities in the Inner Mongolia grassland[J].Journal of Environmental Sciences,2012,24(8):1483-1491.
[99]Zhang N Y,Guo R,Song P,etal.Effects of warming and nitrogen deposition on the coupling mechanism between soil nitrogen and phosphorus in Songnen Meadow Steppe,northeastern China[J].Soil Biology and Biochemistry,2013,65:96-104.
[100]Verena B,Seraina B,Matthias V,etal.Nitrogen deposition effects on subalpine grassland:The role of nutrient limitations and changes in mycorrhizal abundance[J].Acta Oecologica,2012,45:57-65.
[101]Horswill P,O'Sullivan O,Phoenix G K,etal.Base cation depletion,eutrophication and acidification of species-rich grasslands in response to long-term simulated nitrogen deposition[J].Environmental Pollution,2008,155(2):336-349.
[102]Stevensa C J,Duprèc C,Dorland E,etal.The impact of nitrogen deposition on acid grasslands in the Atlantic region of Europe[J].Environmental Pollution,2011,159(10):2243-2250
[103]Liu L L,Greaver T L.A review of nitrogen enrichment effects on three biogenic GHGs:the CO2sink may be largely offset by stimulated N2O and CH4emission[J].Ecology Letters,2009,12(10):1103-1117.
[104]Weier K L,Doran J W,Power J F,etal.Denitrification and dinitrogen/nitrous oxide ratio as affected by soil water,available carbon,and nitrate[J].Soil Science Society of America Journal,1993,57:66-72.
[105]李英臣,宋长春,刘德燕,等.不同氮输入梯度下草甸沼泽土反硝化损失和N2O排放[J].环境科学研究,2009,22(9):1103-1107.
[106]Mosiera A R,Delgadoa J A,Keller M.Methane and nitrous oxide fluxes in Grasslands in western Puerto Rieo[J].Chemosphere,1997,35:2050-2082.
[107]Stephan G,Karl S.Methane and nitrous oxide exchange in differently fertilized grassland in southern Germany[J].Plant and Soil,2001,231:21-35.
[108]Kim Y,Yi M,Koike T.Potential impact of atmospheric N deposition on soil N2O emission varies with different soil N regimes[C].American Geophysical Union,Fall Meeting,2011.
[109]Catchpooir V R,Harper L A,Nyers R J K.Annual Losses of Ammonia from a Grazed Pasture Fertilized with Urea[C].Lexington,Kentucky:Proceedings of the 14th International Grassland Congress,1981:344-347.
[110]Vander Weerden T J,Jarvis S C.Ammonia emission factors for N fertilizers applied to two contrasting grassland soils[J].Environmental Pollution,1997,95:205-211.
[111]张亚欣,范志平,闫加亮,等.氮添加对沙质草地氨挥发及硝态氮淋溶的影响[J].生态学杂志,2011,30(9):1969-1974.
[112]杨江龙.大气CO2与植物氮素营养的关系[J].土壤与环境,2002,11(2):163-166.
[113]Shaw M R,Harte J.Response of nitrogen cycling to simulated climate change:differential responses along a subalpine ecotone[J].Global Change Biology,2001,7(2):193-210.
[114]白洁冰,徐兴良,付刚,等.温度和氮素输入对青藏高原3种高寒草地土壤氮矿化的影响[J].安徽农业科学,2011,39(24):14698-14700,14756.
[115]张璐,黄建辉,白永飞,等.氮素添加对内蒙古羊草草原氮矿化的影响[J].植物生态学报,2009,33(3):563-569.
[116]Gundersen P,Emmett B A,Kjφnaas O J,etal.Impacts of nitrogen deposition on N cycling:a synthesis of NITREX data[J].Forest Ecology and Management,1998,101:37-55.
[117]Haynes R J.Labile organic matter fractions and aggregate stability under short-term,grass-based leys[J].Soil Biology and Biochemistry,1999,31:1821-1830.
[118]Aber J D,McDowell W,Nadelhoffer K,etal.Nitrogen saturation in temperate forest ecosystems:hypotheses revisited[J].BioScience,1998,48:921-934.
猜你喜欢
建材发展导向(2022年24期)2022-12-22
环境卫生工程(2021年4期)2021-10-13
能源工程(2021年3期)2021-08-05
矿产综合利用(2020年1期)2020-07-24
南方周末(2019-06-27)2019-06-27
小学生作文(低年级适用)(2018年12期)2018-03-23
莫愁(2017年15期)2017-05-11
中国老区建设(2016年8期)2016-02-28
植物营养与肥料学报(2011年5期)2011-11-06
植物营养与肥料学报(2011年2期)2011-10-26
