
长白山中华蜜蜂(Apis cerana cerana)遗传多样性分析
2013-12-24 10:00:20于瀛龙周姝婧徐新建朱翔杰周冰峰
福建农林大学学报(自然科学版) 2013年6期
于瀛龙,周姝婧,徐新建,朱翔杰,周冰峰
(福建农林大学蜂学学院,福建福州350002)
遗传多样性是物种适应环境变化的必备条件,是进化的物质基础,反映了物种的进化潜力[1].遗传多样性和遗传结构是种群遗传学、进化生物学和保育遗传学等生命科学的重要内容[1-3].在我国中华蜜蜂遗传多样性和遗传结构的研究中,云南[4]、福建[5-6]、海南[7]、重庆[8]、广东[9]、华东[10]、东北[11]等地区中华蜜蜂的研究已开展[12-13].通过19群样本的测定,在长白山中华蜜蜂遗传多样性的研究中已发现了5种mtDNA单倍型种类[11,14-15];尝试用2群中华蜜蜂样本的csd基因和用12群中华蜜蜂样本的RAPD标记,对长白山中华蜜蜂的遗传多样性和遗传结构进行探索[16-18].长白山地区中华蜜蜂mtDNA标记、微卫星标记的遗传多样性与遗传结构的分析还需深入研究.
长白山是东北中华蜜蜂的主要分布区,是中华蜜蜂分布的东北部边缘,具有独特的生物学特性[19-20].长白山中华蜜蜂在长期的进化过程中形成了群势强、耐寒力高、分蜂性弱等适应当地环境的特性[21],可在最低气温-40℃的环境自然越冬,越冬期长达半年[11,20].长白山中华蜜蜂在体色、体长、前翅翅脉、肘脉指数等主要形态标记表现出特殊性.相比于其他地区中华蜜蜂,长白山中华蜜蜂个体较大,体色偏黑[21-22],肘脉指数较高,为 4.09-5.99[21,23-24],第三径中横脉出现突出的频率较高,为 30%-80%[22,24].
本研究在长白山的腹地,中华蜜蜂主产区采集110群样本,采用5对微卫星引物和mtDNA tRNAleu-COII 357 bp片段,分析该地区中华蜜蜂的遗传多样性与遗传结构.本研究为长白山地区中华蜜蜂资源保护和利用提供重要理论依据.
1 材料与方法
1.1 试验材料
中华蜜蜂样本于2008年8月采自吉林省长白山区的安图县池北区、二道白河镇、松江镇地区,总计样本数量110群蜜蜂(表1).

表1 长白山中华蜜蜂样本信息Table 1 Information of samples from Changbai Mountains
1.2 试验方法
每群取1只工蜂,利用UNIQ10柱式基因组DNA抽提试剂盒(生工生物上海股份有限公司)提取基因组.
PCR扩增微卫星位点的引物来源于NCBI网站(表2),反应条件为95℃预变性5 min;95℃变性50 s,退火30 s(退火温度参照表2),72℃延伸30 s,30个循环;72℃延伸10 min;4℃保存.微卫星PCR产物经8%非变性聚丙烯酰胺凝胶电泳检测.

表2 微卫星位点信息Table 2 Information of 5 microsatellite loci
PCR扩增mtDNA tRNAleu-COII片段,上游引物为E2:5'-GGCAGAATAAGTGCATTG-3',下游引物H2:5'-CAATATCATTGATGACC-3'[25].反应条件为95 ℃预变性5 min;95 ℃变性30 s,50 ℃退火30 s,72 ℃ 延伸30 s,30个循环;72℃延伸8 min;8℃保存.mtDNA PCR产物送至上海英骏生物技术有限公司进行测序.为保证结果的准确性,测序结果中个体数量在3只以下的单倍型进行重复测序.
1.3 统计方法
根据8%非变性聚丙烯酰胺电泳结果进行微卫星位点的等位基因判读,利用Ms tools软件计算期望杂合度(expected heterozygosity,He)、观察杂合度(observed heterozygosity,Ho)、多态信息含量(polymorphism information content,PIC)与等位基因频率.利用Popgene 1.31软件计算等位基因数(observed number of alleles,Na)、有效等位基因数(effective number of alleles,Ne)与香农指数(Shannon's information index,I).
mtDNA测序序列以非编码区“AAAATTT”序列为开始,截取357 bp的序列片段进行分析,利用ClustalX 1.83软件对其进行比对,统计单倍型种类,利用MEGA 4.0软件计算序列的碱基组成,利用DnaSP 5.00.07软件计算单倍型多样度(haplotype diversity,Hd),核苷酸多样度(nucleotide diversity,Pi)与平均核苷酸差异数(average number of nucleotide differences,K).
2 结果与分析
2.1 长白山中华蜜蜂遗传多样性
2.1.1 微卫星位点的遗传多样性 长白山中华蜜蜂在A107、AP043、AP085、AP313、AT101位点整体等位基因较少,遗传多样性水平低.每个微卫星位点发现1-4个等位基因,平均等位基因数为2.6000±1.1402,平均有效等位基因数为1.4181±0.6487.长白山中华蜜蜂5个微卫星位点的期望杂合度为0-0.6107,平均期望杂合度为0.2110±0.2333.除A107位点的期望杂合度为0.6107外,其余4个位点的期望杂合度为0-0.1780.5 个位点的观察杂合度为0-0.1827,平均观察杂合度为0.0645 ±0.0718.5 个位点的平均香农指数为 0.4031 ±0.4202(表3).
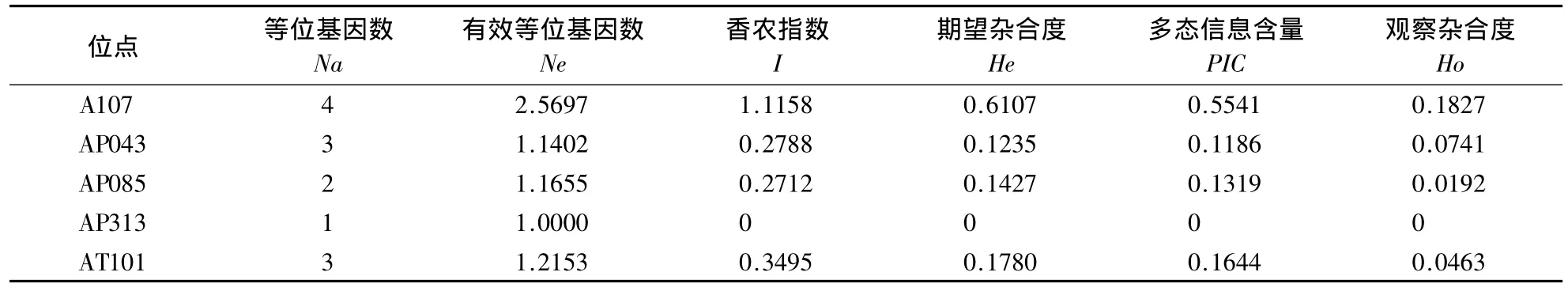
表3 长白山中华蜜蜂微卫星DNA的遗传多样性参数Table 3 Genetic diversity parameters of Apis cerana cerana from Changbai Mountains by microsatellite loci
2.1.2 线粒体DNA遗传多样性 长白山中华蜜蜂在mtDNA tRNAleu-COII 357 bp片段T、C、A、G 4种核苷酸的平均比例分别为44.8%、9.5%、40.6%和5.0%.长白山中华蜜蜂共检测出9种单倍型,H1单倍型(Japan1型,AB078733)是文献报道中分布较为广泛的类型,相比于H1单倍型,1种单倍型在1个位点上发生缺失,7种单倍型分别在1个位点发生突变(表4),其中单一多态位点4个,简约信息位点3个.
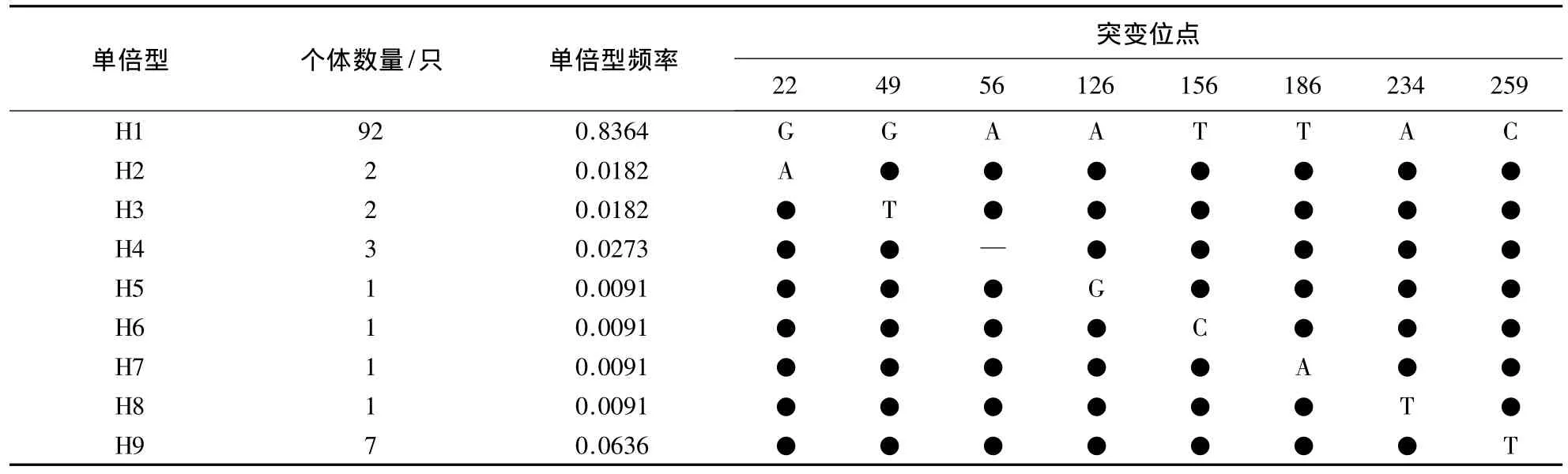
表4 长白山中华蜜蜂mtDNA tRNAleu-COII片段单倍型种类及变异信息1)Table 4 Variable sites and haplotypes of Apis cerana cerana from Changbai Mountains in mtDNA tRNAleu-COII region
长白山中华蜜蜂mtDNA tRNAleu-COII 357 bp片段遗传多样性偏低,单倍型多样度为0.249±0.054、核苷酸多样度为0.00074 ±0.00017、平均核苷酸差异为0.263.
本研究在长白山地区新发现 H2、H3、H4、H5、H7、H8、H9等7种单倍型.与 NCBI网站进行比对,H3(登录号:KF673779)、H5(登录号:KF673782)、H7(登录号:KF673780)、H8(登录号:KF673781)等4种单倍型为中华蜜蜂中首次发现(表4),H2、H4、H9等3种单倍型为在长白山地区发现的新记录.
2.2 长白山中华蜜蜂遗传结构
2.2.1 微卫星遗传结构 长白山中华蜜蜂5个微卫星位点的遗传结构中,均以一种等位基因为主.AP043、AP085、AP313、AT101等4个位点的主要主等位基因,占所检测到的等位基因90%以上.AP313位点只有1种等位基因(342 bp).A107有4种等位基因,其中165 bp占所检测到的等位基因50%以上(表5).2.2.2 线粒体遗传结构 长白山中华蜜蜂mtDNA tRNAleu-COII 357 bp片段的遗传结构较单一.9种单倍型中,以H1(Japan1型)单倍型为主,占83.64%.其次为H9单倍型,占6.36%.其余7种单倍型频率为0.91%-2.73%(表 4).
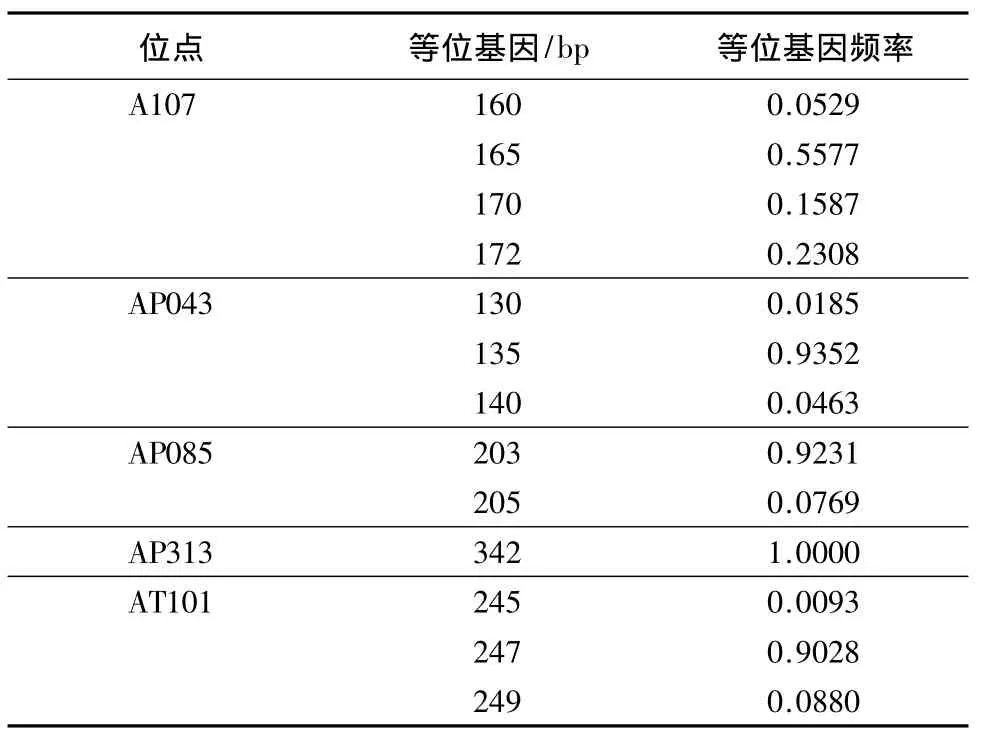
表5 长白山中华蜜蜂的微卫星遗传结构Table 5 Genetic structure of Apis cerana cerana based on microsatellite in Changbai Mountains
3 讨论
(1)在现有中华蜜蜂遗传多样性研究中,中华蜜蜂 A107、AP043、AP085、AP313、AT101 这 5 个微卫星位点的 PIC 值主要范围分别是0.5-0.6、0.2-0.5、0.7-0.85、0.2-0.4 和 0.2-0.5[5-6,26],长白山中华蜜蜂的 PIC 值分别为 0.5541、0.1186、0.1319、0和0.1644,均低于PIC值的主要分布范围或处于较低水平.期望杂合度也体现出相似的规律[5-6,26].目前,较少使用mtDNA tRNAleu-COII片段进行中华蜜蜂遗传多样性研究,仅有海南中华蜜蜂的相关研究中运用了该片段.海南中华蜜蜂 mtDNA tRNAleu-COII片段发现单倍型多样度、核苷酸多样度、平均核苷 酸 差异分别为 0.742 ± 0.017、0.00354 ±0.00013和1.248[7],而长白山中华蜜蜂的这3个指标均远低于海南中华蜜蜂的研究结果.所以微卫星和线粒体分子遗传标记分析,均说明长白山中华蜜蜂遗传多样性水平处于非常低的水平.
(2)长白山植被良好、具有大宗蜜源以及丰富的辅助蜜粉源,是中华蜜蜂优良的栖息地.20世纪前长白山野生中华蜜蜂资源极丰富,此地蜂蜜是皇家贡品[27].由于西方蜜蜂引进后引起种间竞争,导致长白山中华蜜蜂种群数量下降;外来中华蜜蜂基因渗入,使得当地中华蜜蜂面对环境变化、疾病等恶劣条件的耐受性下降;经济较发达的邻国对野生蜂蜜的高价需求,促进毁巢取蜜的行为,毁灭性减少了当地野生中华蜜蜂的种群数量;社会经济的发展,养蜂人的数量减少,养蜂技术下降;人工育王技术应用不当,以上原因均使得长白山中华蜜蜂种群遗传多样性降低.长白山中华蜜蜂形成分散的小种群,遗传变异低于其它地区的中华蜜蜂,并表现出较为单一的遗传结构.
(3)长白山中华蜜蜂是分布在我国东北地区的中华蜜蜂代表,是耐寒型中华蜜蜂的宝贵资源.原长白山中华蜜蜂广泛分布在黑龙江省的小兴安岭以南、吉林省和辽宁省的长白山脉,具有维持强群、耐寒力强等优良蜂种特性.但近年来长白山中华蜜蜂数量大幅度减少,已成为中华蜜蜂中数量最少的生态型,处于濒危维持状态[9,28].长白山中华蜜蜂存在着灭绝的风险.生命资源的灭绝将会永久消失,因此抢救性保护长白山中华蜜蜂刻不容缓.建议在长白山中华蜜蜂分布较多、生态条件较好的地方设立中华蜜蜂保护区;在保护区内政府用补贴、技术支持等形式鼓励当地农民原始饲养本土的中华蜜蜂,扩大长白山中华蜜蜂的种群数量;禁止西方蜜蜂和转地的中华蜜蜂进入,禁止从保护区外购买蜂种或蜂王,保持长白山中华蜜蜂的遗传特征;在保护区内适当的有计划的蜂群交流,不进行人工选育,保持长白山中华蜜蜂的遗传多样性.致谢:样本采集中得到了杨明福等同志的帮助.本科生夏凤枝参与部分实验.谨此致谢!
[1]法兰克汉,巴卢,布里斯科.保育遗传学导论[M].北京:科学出版社,2005:30.
[2]沈银柱,黄占景,王正询,等.进化生物学[M].北京:高等教育出版社,2008:112-113.
[3]徐晋麟,赵耕春.基础遗传学[M].北京:高等教育出版社,2009:282-284.
[4]殷玲,吉挺,陈国宏.利用微卫星标记分析云南省6个东方蜜蜂(Apis cerena)群体遗传多样性及遗传分化[J].西南农业学报,2011,24(2):772-778.
[5]朱翔杰,徐新建,周妹婧,等.武夷山自然保护区中华蜜蜂微卫星DNA遗传分析[J].福建农业学报,2011,26(6):935-940.
[6]朱翔杰,周冰峰,吴显达,等.福建中华蜜蜂微卫星标记的遗传多样性分析[J].福建农林大学学报:自然科学版,2011,40(4):407-411.
[7]周姝婧,徐新建,朱翔杰,等.海南中华蜜蜂线粒体DNA的遗传多样性[J].福建农林大学学报:自然科学版,2012,41(2):170-175.
[8]曹联飞,姬聪慧,任勤,等.重庆金佛山地区东方蜜蜂遗传多样性研究[J].西南师范大学学报,2010,35(5):162-166.
[9]成纯富,周丹银,刘意秋,等.基于mtDNA Cytb基因序列分析广东省东方蜜蜂遗传多样性[J].蜜蜂杂志,2012(9):4-6.
[10]吉挺,殷玲,刘敏,等.华东地区中华蜜蜂六地理种群的遗传多样性及遗传分化[J].昆虫学报,2009,52(4):413-419.
[11]李志勇,薛运波,赵惠燕.长白山区中华蜜蜂mtDNA非编码区至COⅠ基因PCR及序列分析[J].湖北农业科学,2009,48(5):1051-1054.
[12]YIN L,JI T,LIU M,et al.Genetic diversity and relationship between genetic distance and geographical distance of 6 Apis cerana cerana populations in China[J].Research Journal of Animal Sciences,2008,2(6):183-187.
[13]SMITH D R,VILLAFUERTE L,OTIS G,et al.Biogeography of Apis cerana F.and A.nigrocincta:insight from mtDNA studies[J].Apidologie,2000,31(2):265-279.
[14]TAN K,WARRIT N,SMITH D R.Mitochondrial DNA diversity of Chinese Apis cerana[J].Apidologie,2007,38(3):238-246.
[15]姜玉锁,赵慧婷,姜俊兵,等.中国境内不同地理型东方蜜蜂线粒体tRNAleu-COII基因多态性研究[J].中国农业科学,2007,40(7):1532-1542.
[16]刘志勇,王子龙,王欢,等.中华蜜蜂csd多态性分析[J].中国农业科学,2011,44(23):4911-4917.
[17]薛运波,李兴安,葛凤晨,等.长白山中华蜜蜂基因组DNA多态性的研究[J].中国农业科学,2007(40):426-432.
[18]薛运波,李兴安,葛凤展,等.长白山中华蜜蜂基因组CBS-700(bp)DNA同源片段克隆及其Southern杂交的研究[J].中国蜂业,2006,57(10):8-11,17.
[19]杨冠煌.中华蜜蜂[M].北京:中国农业科技出版社,2001:13.
[20]龚一飞,张其康.蜜蜂分类与进化[M].福建:科学技术出版社,2000:21.
[21]杨冠煌,李桂仙.中华蜜蜂分布和变异(五)—东北地区中华蜜蜂资源[J].中国养蜂,1983(5):19.
[22]杨冠煌,许少玉,匡邦郁,等.东方蜜蜂Apis cerana Fab在我国的分布及其亚种分化[J].云南农业大学学报,1986,12(1):89-92.
[23]张连江,樊贤,谭垦,等.长白山东方蜜蜂形态特征研究[J].云南农业大学学报,2006,21(4):511-516.
[24]葛凤晨,历延芳,柏建民,等.长白山中华蜜蜂分布及其生产效率的研究[J].中国养蜂,2002,53(6):4-7.
[25]GARNERY L,COMUET J M,SOLIGANAC M.Evolutionary history of the honey bee Apis mellifera inferred from mitochondrial DNA analysis[J].Moleculor Ecology,1992,1(3):145-154.
[26]徐新建,周姝婧,朱翔杰,等.海南岛中华蜜蜂遗传多样性的微卫星DNA分析[J].昆虫学报,2013,56(5):554-560.
[27]葛凤晨,陈东海,历延芳,等.长白蜜蜂文化研究[M].长春:吉林科学技术出版社,2006:1-21.
[28]国家畜禽遗传资源委员.中国畜禽遗传资源志:蜜蜂志[M].北京:中国农业出版社,2011:45-49.
猜你喜欢
特产研究(2022年6期)2023-01-17 05:05:06
种子(2021年3期)2021-04-12 01:42:22
学生天地(2020年34期)2020-06-09 05:50:38
辽金历史与考古(2019年0期)2020-01-06 07:44:54
四川动物(2017年4期)2017-07-31 23:54:19
外语教学理论与实践(2016年1期)2016-06-11 05:51:48
集美大学学报(自然科学版)(2015年4期)2015-02-28 01:13:37
中国工程咨询(2015年8期)2015-02-16 06:39:04
河北遥感(2014年3期)2014-07-10 13:16:48
华东理工大学学报(自然科学版)(2014年1期)2014-02-27 13:48:29
