
GST-NDPK-A融合蛋白的原核表达及体外互作蛋白的检测
2013-12-23 05:12吕芬刘忠银兴峰赵振岭陈妙娟张嘉萱陈伟钱垂文熊盛
生物技术通报 2013年5期
吕芬 刘忠 银兴峰 赵振岭 陈妙娟 张嘉萱 陈伟 钱垂文 熊盛
(1.暨南大学生命科学技术学院 生物医药研究开发基地,广州 510632;2.暨南大学生命科学技术学院 生命与健康工程研究院,广州 510632)
核苷二磷酸激酶A(nucleoside diphosphate kinase A,NDPK-A),是Nm23-H1基因编码的多功能蛋白,可以调节细胞增殖、分化、发育、迁移、凋亡,信号转导等多种生物学活动[1]。Nm23-H1最早被鉴定为肿瘤转移抑制基因以来,一直是肿瘤研究的热点。针对多种肿瘤细胞株,如黑色素瘤、乳腺癌、前列腺癌、结肠癌、肝癌等,Nm23-H1的过表达表现出抑制细胞迁移的能力;然而,在恶性白血病、神经母细胞瘤、骨肉瘤患者体内,Nm23-H1的高表达却使生存率降低;在鼻咽癌、直肠癌中,Nm23-H1的表达与肿瘤转移并无明显关系[2]。这种复杂的调节模式使得Nm23-H1成为决定肿瘤细胞命运的关键分子,而成为备受关注的药物作用靶标。关于Nm23-H1多功能调节的分子机制,目前还未被透彻地揭示。众所周知,肿瘤的发生、转移的分子基础是一个立体、整体水平上的蛋白质间相互作用的结果。目前,越来越多的研究聚焦于去发现在重要的信号转导途径中,与NDPK-A相互作用的伙伴蛋白。如GTP结合蛋白[3]、转录调控因子[4]、SET复合物[5]等以及与相关蛋白结合参与信号转导,如Erk Map激酶途径[6]、TGF-β信号途径[7]等,证据表明,NDPK-A与这些蛋白相互作用、相互影响、共同调控,涉及肿瘤细胞的发生、增殖、迁移、凋亡、分化、血管生成、DNA修复等各个方面的相关性。为了进一步研究NDPK-A在细胞中扮演的角色,本研究通过基因工程技术体外构建表达载体,诱导可溶性GST-NDPK-A融合蛋白的高效表达,亲和纯化,利用GST-Pull down技术在肺腺癌A549细胞中筛选与NDPK-A结合的蛋白质,并经SDS-PAGE电泳分离,LC-MS/MS质谱分析进行鉴定,试图寻找与NDPK-A相互作用的靶蛋白,旨在为研究NDPK-A的功能和作用机制提供一个新的切入点。
1 材料与方法
1.1 材料
1.1.1 菌种、质粒和细胞系 大肠杆菌(Escherichia coli)DH5α、BL21(DE3)为本实验室保存、质粒pBV-NDPK-A由本实验室构建并保存;pGEX-4T-2质粒购自上海希匹吉生物公司;肺腺癌细胞株A549由本实验室储备。
1.1.2 主要试剂 限制性内切酶BamHⅠ、EcoRⅠ、T4 DNA连接酶均购自TaKaRa公司;DNA凝胶回收试剂盒、质粒提取试剂盒均购自Omega公司;KOD-Plus-Neo酶购自TOYOBO公司;Glutathione Sepharose 4B凝胶树脂购自GE Healthcare公司;氨苄青霉素、IPTG、PMSF均购自Sigma公司;完全型蛋白酶抑制剂CPI购自Roche公司;NP40细胞裂解液、BCA试剂盒均购自碧云天生物公司;NDPK-A鼠单抗购自Santa Cruz公司;5 cm分拆式层析柱购自Bio-Rad公司;PCR引物由上海生工生物公司合成。
1.2 方法
1.2.1 重组质粒pGEX-4T-NDPK-A的构建 根据GenBank检索到的人源NDPK-A全基因序列以及pGEX-4T-2空载的多克隆位点图谱设计引物。上游引物为:5'-CGGGATCCATGGCCAACTGTGAGCGTA-3',下划线为BamH I酶切位点;下游引物:5'-AGCCGG AATTCTCATTCATAGATCCAGTTCTGAGC-3',下划线为EcoR I酶切位点。以含有人源全长目的基因的质粒pBV-NDPK-A为模板,进行PCR扩增,条件为:94℃ 2 min;98℃ 10 s,66℃ 30 s,68℃ 30 s,30个循环;68℃ 5 min。将PCR产物克隆入pGEX-4T-2载体中,PCR及双酶切鉴定后,交由上海英骏生物公司广州分公司测序。
1.2.2 GST-NDPK-A融合蛋白的诱导表达 将测序正确的GST-NDPK-A重组质粒转化至大肠杆菌BL21感受态细胞中,在含氨苄青霉素抗性的固体LB培养基上37℃倒置培养过夜,挑取阳性单克隆,接种于5 mL含氨苄青霉素(100 μg/mL)的LB液体培养基中,37℃振荡(180 r/min)活化培养过夜。为了尽可能获得足量的可溶性蛋白用于后续试验,将逐级培养活化后的菌液按1∶100的比例接种于300 mL含氨苄青霉素的LB液体培养基中,培养至OD600达到0.6-0.8时,加入IPTG至终浓度为0.2 mmol/L,20℃下100 r/min继续诱导培养过夜。
1.2.3 GST-NDPK-A融合蛋白的纯化、鉴定 离心收集上述诱导菌体,按10∶1(V/V)的比例重悬于 裂 解 缓 冲 液[50 mmol/L Tris-HCl、pH7.6、150 mmol/L NaCl、2 mmol/L EDTA、1% NP40(V/V),及蛋白酶抑制剂CPI和PMSF]中,并加入终浓度为1 mg/mL的溶菌酶,涡旋混匀,冰浴30 min;冰浴超声裂解15 min后,将破碎液于4℃,14 000 r/min,离心30 min,收集上清。吸取0.5 mL Glutathione Sepharose 4B凝胶珠子混悬液至5 cm的小层析柱内,静置30 min;沉降完全后,加入6倍柱体积的EBC缓冲液清洗平衡纯化柱;然后,将收集的细菌裂解液上清过柱;通过使用不同盐浓度梯度的washing buffer 洗脱杂蛋白;经EBC缓冲液平衡柱子后,收集偶联有目的融合蛋白的凝胶珠子于0.5 mL的EBC缓冲液中,并直接储存于分拆式小层析柱内。取适量凝胶珠子溶于2×SDS-PAGE buffer中,100℃加热变性,离心取上清,进行凝胶电泳分析。并以NDPK-A鼠单抗为一抗,鼠源IgG为二抗进行Western blot验证。
1.2.4 GST-Pull down试验 在8个100 mm培养皿中培养A549细胞,待单层细胞长至约95%融合时,用细胞刮收集细胞于2 mL含有蛋白酶抑制剂的NP40裂解缓冲液中,混匀冰浴裂解40 min,每间隔10 min震动一次。裂解完全后,13 200 r/min,4℃离心30 min,收集上清,即为细胞总蛋白(全细胞裂解液)。BCA法测定蛋白浓度后,立即用于GST- Pull down试验。
取含GST蛋白的纯化珠子与全细胞裂解液在4℃旋转孵育1 h,离心后,转移上清至新的离心管中,即为预吸附后的全细胞裂解液。将其均分为2份,一份加入含融合蛋白的纯化珠子,另一份加入含GST的纯化珠子,根据BCA法测定的蛋白浓度,调整加入体积,确保二者珠子上结合的总蛋白量一致,在4℃下旋转孵育过夜。之后,用预冷的washing buffer温和清洗珠子上非特异性结合的杂蛋白,重复4次,经SDS-PAGE电泳后,银染分析。
1.2.5 差异蛋白条带的酶解及质谱鉴定 数次银染分析后,鉴定出有重复性差异的蛋白条带,继而将蛋白样品扩大上样量,进行SDS-PAGE电泳,考马斯亮蓝染色后,从SDS-PAGE胶上切取差异条带,经过常规的脱色、还原、烷基化、胰酶酶解、肽段萃取等处理,后续样本冻干、溶解、Trap柱脱盐及LC-MS/MS质谱分析。
2 结果
2.1 重组质粒pGEX-4T-NDPK-A的构建
以pBV-NDPK-A为模板进行PCR扩增,获得分子量约500 bp的目的片段(图1)。将PCR扩增获得的全长NDPK-A克隆至原核表达载体pGEX-4T-2中,构建pGEX-4T-NDPK-A重组质粒。挑选阳性单克隆,重组质粒提取后,进行PCR鉴定和双酶切鉴定(图2)。PCR鉴定获得一特异性的目的条带,双酶切结果与预期结果吻合。测序结果表明目的片段序列完整且按通读框方式插入pGEX-4T-2载体中。

图1 目的基因的PCR扩增

图2 重组表达质粒的鉴定
2.2 GST-NDPK-A融合蛋白的表达、纯化
如图3所示,已成功对可溶性上清分布的融合蛋白GST-NDPK-A、GST蛋白进行诱导表达纯化,纯度达到90%以上。
2.3 GST-NDPK-A融合蛋白的鉴定
如图4所示,融合蛋白GST-NDPK-A能发生特异性的抗原抗体反应,具有免疫学特异性。
2.4 GST-Pull down筛选及复合蛋白的SDS-PAGE分离
以GST-NDPK-A融合蛋白为诱饵蛋白从A549全细胞裂解液中捕获的蛋白质经SDS-PAGE电泳后,银染与考马斯亮蓝染色的结果(图5)对应显示,与对照组蛋白带比较,发现在25 kD左右处存在明显的差异蛋白带,即在A549细胞中找到与NDPK-A潜在性特异结合和相互作用的蛋白。切取考马斯亮蓝染色的差异蛋白带并将对照GST蛋白Pull down组中相应位置的胶也切下,一起进行LC-MS/MS鉴定。

图3 GST-NDPK-A蛋白(A)和GST蛋白(B)诱导表达及纯化

图4 融合蛋白GST-NDPK-A的Western blot鉴定
2.5 NDPK-A相互作用蛋白的质谱鉴定
LC-MS/MS鉴定出的初级数据处理后用Mascot搜索引擎进行IPI human protein database(V3.68)搜库。结果(图6,表1)分析显示,Fussel-18(functional Smad-suppressing element on chromosome 18)和Rrp12(ribosomal RNA processing 12)成为NDPK-A潜在的体外相互作用蛋白。

图5 GST-Pull down银染分析(A)及考染分析(B)
3 讨论
寻找互作蛋白能够为深入揭示NDPK-A在细胞中的功能提供新的依据和线索。GST-Pull down技术是体外确定两个已知蛋白之间是否存在相互作用以及筛选与已知融合蛋白相互作用的未知蛋白的有效方法。因此,本研究构建GST-NDPK-A融合蛋白表达载体,以最优条件诱导获得部分可溶性GSTNDPK-A融合蛋白,上清过柱后,亲和固化在谷胱甘肽琼脂糖珠上,此为纯化后的诱饵蛋白。而前期经Western blot验证NDPK-A蛋白在肺腺癌A549细胞系中高表达,说明可在此细胞系中寻找NDPK-A的相互作用蛋白。诱饵蛋白与A549细胞裂解液以最佳时间孵育后,捕捉到细胞中有特异性相互作用的目的靶蛋白,最后GST融合蛋白-目的蛋白复合物一起被谷胱甘肽琼脂糖珠沉降下来,沉淀的复合物经过SDS-PAGE电泳分离,从电泳结果发现,在25 kD左右处出现新的条带,说明在A549中成功筛选到了NDPK-A潜在的相互作用蛋白。
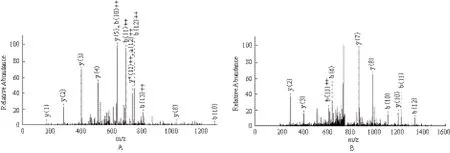
图6 用LC-MS/MS质谱鉴定SDS-PAGE分离的差异蛋白条带

表1 结合蛋白的分类
目前蛋白质的鉴定常用的有两种质谱分析技术[8]:其中,MALDI-TOF-MS鉴定蛋白质是依靠肽质量指纹图(PMF)来实现,要求待分析的蛋白质样品纯度较高,而SDS-PAGE分离的蛋白质带通常含有多个蛋白质。并且,对于蛋白质分子普遍存在的翻译后修饰,如多肽侧链上的糖基化、磷酸化、乙酰化等化学修饰,MALDI-TOF-MS无法鉴定,不能获得多肽序列,造成鉴定结果准确性不够。而本研究采用LC-MS/MS鉴定蛋白质,通过液相色谱分离多肽片段,样品不纯对分析影响较小,不仅可以准确测定蛋白质和肽段的分子量,还能获得肽段的序列标记及其侧链化学修饰等数据,提高了蛋白质鉴定结果的准确性。此外,我们不仅对GSTNDPK-A融合蛋白Pull down的差异蛋白条带进行了质谱鉴定,而且对相应位置处GST标签Pull down的蛋白亦做了鉴定,以GST标签Pull down的质谱结果为对照,删除了一些可能与GST-NDPK-A融合蛋白中的GST标签结合的互作蛋白,如:Glutathione S-transferase P,通过试验过程中GST标签蛋白的预吸附和质谱鉴定结果的对照设定,大大提高了NDPK-A互作蛋白筛选的准确性。
质谱鉴定出两个可能与NDPK-A结合的相互作用蛋白,分别是Fussel-18(functional Smad-suppressing element on chromosome 18)和Rrp12(ribosomal RNA processing 12),两者均为文献中之前未见报道的新的NDPK-A结合蛋白。NDPK-A广泛的分布于细胞浆中,少量存在于细胞核、细胞膜、线粒体、微管和组织液中[9],其功能的发挥与它在细胞中的定位关系密切。鉴定到的Fussel-18主要定位于细胞核与细胞质,而Rrp12表达于细胞核,这与NDPK-A的分布是相一致的。这说明重组融合蛋白GST-NDPK-A是有功能的,并且适合于招募细胞中存在的相互作用蛋白伙伴。
Fussel-18,属于癌蛋白Ski家族[10]。研究发现,在神经系统中,Fussel-18与Smad2和Smad3蛋白相互作用,抑制了TGF-β的信号转导[10]。TGF-β信号的传递过程由Smad家族蛋白参与调控,并与细胞的生长、增殖、分化、凋亡相关[11,12]。有报道显示,TGF-β受体的相互作用蛋白之一STRAP(丝苏氨酸激酶受体相关蛋白),它通过促进TGF-β受体与Smad7蛋白作用的稳定性,实现对TGF-β信号传递的抑制[13]。另有研究显示,NDPK-A与STRAP可以利用半胱氨酸残基上二硫键交联的方式,直接结合互相作用,不仅能够对STRAP介导的TGF-β信号的抑制作用起到增强效应,还能对TGF-β调节的凋亡和生长带来影响[7]。这里暗示NDPK-A通过与STRAP的直接结合,可能成为TGF-β信号的负调控者。此外,TGF-β在癌症生物学中被认为能够抑制正常细胞的增殖,反而促进转移性肿瘤细胞的增殖[14]。这些结果引导我们提出疑问:NDPK-A介导调节TGF-β信号是否与NDPK-A肿瘤转移抑制功能相关。因此,进一步研究NDPK-A与Fussel-18的相互作用关系,预期能更深层次地揭示NDPK-A调节TGF-β信号的分子机制及与肿瘤抑制相关的生物学功能。
本研究鉴定到的另一个蛋白Rrp12,作为前体40S和60S亚基的一个稳定组成部分,参与调节核糖体亚基成熟与出口[15]。这暗示着NDPK-A在细胞内可能与核糖体组装有关。近来研究发现,Rrp12影响着细胞周期的进程,Rrp12的缺失会导致S期进入障碍、S期以及有丝分裂期的延滞[16]。随着对细胞周期研究的不断加深,人们发现细胞周期紊乱与肿瘤的发生有密切关系,并提出“肿瘤可能是一类细胞周期性疾病”。前期研究发现,NDPK-A可以使宫颈癌细胞Caski和SiHa细胞周期延滞在G1期,并有效抑制细胞增殖[17]。此外,也有报道,Rrp12参与了DNA损伤反应的调节[16]。在NDPK-A众多生物学活性中,3'-5'核酸外切酶活性,与其具备的DNA损伤修复功能密切相关[18]。NDPK-A的低表达削弱了DNA损伤修复功能,基因发生突变,染色体异常,随之会发生细胞癌变。因此,验证Rrp12与NDPK-A的相互作用关系,探讨这二者在细胞周期及DNA损伤反应中的机理与意义,有助于研究它们在肿瘤发展过程中的生理功能。
通过初步的试验结果可知,NDPK-A与Fussel-18及Rrp12在体外可能存在相互作用,但是NDPK-A与这二者在体内外的相互作用情况、及具体的结合区段,还需要进一步的研究确证,它们相互作用的生物学功能及意义也有待阐明,这将为基于NDPK-A的药靶研究增加可靠的试验及理论依据。
4 结论
本研究结合经典分子生物学与蛋白组学方法,通过构建融合蛋白表达载体、高效诱导可溶性表达、与谷胱甘肽琼脂糖珠亲和层析,实现了GSTNDPK-A的原核表达和纯化,并采用GST-Pull down联合LC-MS/MS质谱分析技术筛选鉴定到两个与NDPK-A的互作蛋白,分别为Fussel-18、Rrp12。
[1] Narayanan R, Ramaswami M. Regulation of dynamin by nucleoside diphosphate kinase [J]. J Bioenerg Biomembr, 2003, 35(1):49-55.
[2] Marino N, Marshall JC, Steeg PS. Protein-protein interactions:a mechanism regulating the anti-metastatic properties of Nm23-H1 [J]. Naunyn-Schmiedeberg’s Arch Pharmacol, 2011, 384(6):351-362.
[3] Tseng YH, Vicent D, Zhu J, et al. Regulation of growth and tumorigenicity of breast cancer cells by the low molecular weight GTPase Rad and nm23 [J]. Cancer Research, 2001, 61(5):2071-2079.
[4] Kaul R, Murakami M, Lan K, et al. EBNA3C can modulate the activities of the transcription factor Necdin in association with the metastasis suppressor protein Nm23-H1 [J]. Journal of Virology, 2009, 83(10):4871-4883.
[5] Fan Z, Beresford PJ, Oh DY, et al. Tumor suppressor NM23-H1 is a granzyme A-activated DNase during CTL-mediated apoptosis, and the nucleosome assembly protein SET is its inhibitor [J]. Cell, 2003, 112(5):659-672.
[6] Salerno M, Palmieri D, Bouadis A, et al. Nm23-H1 metastasis suppressor expression level influences the binding properties, stability and function of the Kinase Suppressor of Ras (KSR1) Erk scaffold in breast carcinoma cells [J]. Molecular and Cellular Biology, 2005, 25(4):1379-1388.
[7] Seong HA, Jung H, Ha H. NM23-H1 tumor suppressor physically interacts with serine-threonine kinase receptor-associated protein, a transforming growth factor-beta (TGF-beta)receptor-interacting protein, and negatively regulates TGF-beta signaling [J]. Journal of Biological Chemistry, 2007, 282(16):12075-12096.
[8] 詹显全, 陈主初. 蛋白质组中蛋白质鉴定技术的研究近况. 国外医学:分子生物学分册, 2002, 24 (3):129-130.
[9] Urano T, Furukawa K, Shiku H. Expression of nm23/NDP kinase proteins on the cell surface [J]. Oncogene, 1993, 8(5):1371-1376.
[10] Arndt S, Poser I, Schubert T, et al. Cloning and functional characterization of a new Ski homolog, Fussel-18, sepecifically expressed in neuronal tissues [J]. Laboratory Investigation, 2005, 85(10):1330-1341.
[11] Heldin CH, Miyazono K, Ten Dijke P. TGF-beta signalling from cell membrane to nucleus through SMAD proteins [J]. Nature, 1997, 390(8):465-471.
[12] Elliott RL, Blobe GC. Role of transforming growth factor beta in human cancer [J]. Journal of Clinical Oncology, 2005, 23(9):2078-2093.
[13] Seong HA, Jung H, Choi HS, et al. Regulation of transforming growth factor-β signaling and PDK1 kinase activity by physical interaction between PDK1 and serine-threonine kinase receptorassociated protein [J]. Journal of Biological Chemistry, 2005, 280(52):42897-42908.
[14] Bierie B, Moses HL. TGF-beta and cancer [J]. Cytokine Growth Factor Rev, 2006, 17(2):29-40.
[15] Oeffinger M, Dlakic M, Tollervey D. A pre-ribosome-associated HEAT-repeat protein is required for export of both ribosomal subunits [J]. Genes Dev, 2004, 18:196-209.
[16] Dosil M. Ribosome synthesis-unrelated functions of the preribosomal factor Rrp12 in cell cycle progression and the DNA damage response [J]. Molecular and Cellular Biology, 2011, 31(12):2422-2438.
[17] 杨琰, 卢实, 李敏芳, 等.肿瘤转移抑制基因nm23-H1对不同宫颈癌细胞侵袭和增殖的作用[J].癌症, 2009, 28(7):702-707.
[18] Ma D, McCorkle JR, Kaetzel DM. The metastasis suppressor Nm23-H1 possesses 3’-5’ exonuclease activity [J]. Journal of Biological Chemistry, 2004, 279(17):18073-18084.
猜你喜欢
肝博士(2022年3期)2022-06-30
海外星云(2021年9期)2021-10-14
食品安全导刊(2021年20期)2021-08-30
食品科学(2018年10期)2018-05-23
当代化工研究(2016年5期)2016-03-20
西南医科大学学报(2015年1期)2015-08-22
中国当代医药(2015年9期)2015-03-01
西南军医(2015年6期)2015-01-23
特产研究(2014年4期)2014-04-10
