
黑龙江省药用植物根际土壤真菌多样性
2013-12-20 02:20慕东艳吕国忠孙晓东赵志慧
生态学报 2013年1期
慕东艳,吕国忠 ,孙晓东,王 娜,赵志慧
(1.辽宁师范大学生命科学学院,大连 116029;2.大连民族学院环境与资源学院,大连 116600)
植物根际土壤中通常生长着大量各种类型的微生物,包括细菌、真菌、放线菌等[1]。其中许多微生物能通过固氮作用,促进磷等矿物质营养的吸收以及抑制土传病原物等方式促进植物的生长和发育,保护植物健康生长[2]。但是,某些微生物却以植物根系作为侵染点侵染植物,从而对植物的生长造成危害,包括某些致病真菌、卵菌、细菌和线虫等。这些土传致病菌在时间和空间上的传播对于植物的生长造成了极大的危害。另外,植物根际土壤中各种有益微生物通过竞争、拮抗与重寄生等作用有效地抑制了土传病害的发生与传播。植物根际有益微生物和有害微生物之间的生态关系对于植物健康生长与发育发挥着巨大的不可替代的作用[3-5]。
黑龙江省是我国中药材主要产区之一,其中药材的产销总量占到全国的15%左右,其所产北药也因其质量好而畅销国内外。特别是五味子、平贝母和柴胡的人工栽培地区广泛,栽培面积大,产量高。近年来,随着药用植物种植面积的增加以及连作或复种面积的不断扩大,随之而产生的土传病害也越来越严重,已经成为生产中的难点问题[6]。大多数名贵中药材的药用部位是根及根茎等地下部分,而土传病原菌则侵染和危害中药材的根部,引起根部病害,严重影响中药材的质量和产量,并造成重要经济损失[7-8]。
随着药用植物病害危害的不断加重,药农使用化学农药的种类和剂量不断增加,不仅污染了环境,而且造成中药材农药残留量严重超标,严重影响药材质量,不利于药材的加工甚至出口。利用拮抗真菌防治药用植物土传病害不仅是生产无公害和优质中药材的保证,同时也能够保持生态平衡,实现农业可持续发展[9]。由于不同的药用植物或同一药用植物的不同生长期其根系分泌物不同,其根际微生物种类、数量和优势菌种均不同。研究药用植物根际土壤真菌多样性,从中可以筛选到更多拮抗真菌或有益真菌,为研制生防菌剂和开展药用植物根际土传病害的生物防治提供生防菌种资源保障。另外,在药用植物根际土壤中蕴藏着大量具有生物转化能力的真菌种类,可用于中药活性成分的生物转化[10]。因此,研究不同的药用植物根际土壤真菌的多样性具有重要的理论研究和实际应用价值。
本研究采用传统的形态学方法对分离自黑龙江省多种药用植物根际土壤的真菌进行鉴定,并对其种群组成和生态分布进行统计与分析。本文研究的首要目的是要明确黑龙江省药用植物根际土壤真菌的种类资源及区系分布特点,同时为筛选针对药用植物土传病害的拮抗生防菌株和具有生物转化功能的菌株提供丰富真菌资源。
1 材料与方法
1.1 土壤样品的采集
在2010年7月份和10月份,对黑龙江省伊春、铁力、绥化、哈尔滨、牡丹江、佳木斯等6个地区栽培的药用植物的根际土壤进行取样,共采集土壤样品220份(表1),采集的药用植物种类有五味子、平贝母、刺五加、防风、党参、柴胡、黄芩、桔梗、穿山龙、牛蒡、知母、射干、益母草和红花,共14种。采样方法为三点或五点混合采样法。土壤样品为采自粘附在不同药用植物根系上和受根系分泌物影响的土壤颗粒,即根际土壤。在某一地区某种药用植物种植区内选取3个或5个(根据种植面积来选取)采集点,采样前除去药用植物主根系周围土壤表层的枯枝落叶等杂物后,将表层2—3 cm的土层(即未粘附在主根系上的土壤)铲去,然后选取距植物主根系表面0—1 cm范围内,沿主根系2—15 cm深处土层范围内的土壤样品(即受植物根系分泌物影响的土壤)进行采集,多点采集的土壤样品经充分混合后取约100 g土壤装入无菌的塑料袋中,并记录采样时间、地点和采集样品种类,带回实验室后立即进行真菌的分离。

表1 黑龙江省药用植物根际土壤样品的采集地点Table 1 The rhizosphere soil sampling regions of medicinal plants grown in Heilongjiang Province
1.2 土壤真菌的分离
采用稀释平板法和土壤颗粒平板法进行土壤真菌的分离,分离培养基为孟加拉红培养基。(1)稀释平板法:称取10 g土壤样品放入装有90 mL无菌水的三角瓶中,将三角瓶置于摇床上振荡25—30min,使土壤颗粒均匀分散于无菌水中,得到稀释倍数为10的土壤悬浮液,从中吸取1 mL移入装有9 mL无菌水的试管中,充分振荡混匀,为稀释倍数102的悬浮液,再吸取10-2稀释液1 mL,移入装有9 mL无菌水的试管中,混匀后即成稀释倍数为103的悬浮液;依此类推,制成10-4的土壤稀释液。吸取10-2、10-3、10-4稀释液1mL置于培养皿中,每个稀释度重复2次,与已熔化并冷却至40℃左右的孟加拉红培养基混合均匀,然后置于25℃恒温培养箱中培养。(2)土壤颗粒平板法:用灭菌的小扁勺取土样0.005 g左右,分散、弄碎后,将其均匀散布在凝固了的孟加拉红培养基上,每个土样重复2次,然后将培养皿置于25℃恒温培养箱内培养。5—7 d后,对分离到的真菌菌落进行计数和分析,将单个菌落挑出纯化并保存菌种。
1.3 土壤真菌的纯化与鉴定
通过尖端菌丝挑取法,将在孟加拉红平板培养基上生长出的不同形态和颜色的真菌菌落转接入PDA培养基或其它标准培养基(SNA、SMA、CYA、CA、MEA、PCA培养基等)上进行纯化培养,于25℃恒温培养箱中培养5—15 d后,在光学显微镜下进行显微形态观察,参照相关真菌分类文献[11-16]进行真菌形态鉴定。
1.4 数据处理
分离频率用于比较和判断真菌的优势种群,分离频率的计算公式:
分离频率(%)=某一分离物的总数(株)/所有分离物总数(株)×100%
真菌种群的多样性水平通过Shannon多样性指数(H')、Pielou均匀度指数(J)和Margalef丰富度指数(R)进行分析:
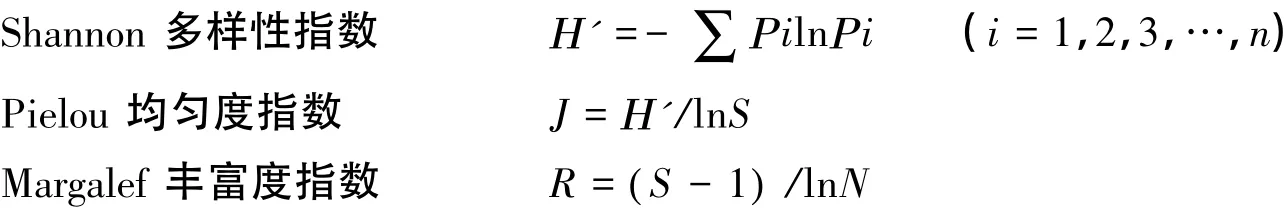
式中,Pi是第i种的个体数占总个体数的比例,可以用Ρi=Ni/N求出,Ni为第i种物种个体数,N为总个体数,S为每个样品中的物种总数。
Jaccard相似性指数

式中,a,b分别为两种生境类型中真菌的种数或属数,c为两种生境类型中共有的真菌种数或属数,用于比较两个生境中真菌种类组成的相似程度。根据Jaccard相似性系数原理:当Cj为0.00—0.25时,为极不相似;当Cj为0.25—0.50 时,为中等不相似;当Cj为 0.50—0.75 时,为中等相似;当Cj为0.75—1.00 时,为极相似。
2 结果与分析
2.1 黑龙江省药用植物根际土壤真菌的种类与数量组成特点
通过对黑龙江省6个地区的14种药用植物根际土壤真菌的分离培养,共分离得到真菌1016株,其中805株真菌经形态鉴定明确到种级分类单位,它们属于35属86种(表2)。而其它211株为培养条件下不产生子实体结构而无法明确其分类地位的真菌,统一归入无孢类群,根据菌落及菌丝形态特征,将它们初步归为5个类群组。试验中经分离和鉴定出的真菌种类分别是接合菌门的被孢霉属Mortierella、毛霉属Mucor、小克银汉霉属Cunninghamella、犁头霉属Absidia、孢球托霉属Gongronella、根霉属Rhizopus、接霉属Zygorhynchus共7属10种,占总菌株数的7.78%;子囊菌门的毛壳属Chaetomium,即1属2种,占0.69%;无性型真菌的27属74种,占70.76%,分别是青霉属Penicillium14个种,曲霉属Aspergillus12个种,木霉属Trichoderma8个种,镰孢菌属Fusarium5个种,金孢属Chrysosporium4个种,轮枝菌属Verticillium3个种,枝顶孢属Acremonium、腐质霉属Humicola、枝孢属Cladosporium、粘帚霉属Gliocladium、漆斑菌属Myrothecium、拟青霉属Paecilomyces、帚霉属Scopulariopsis各2个种,链格孢属Alternaria、节菱孢属Arthrinium、白僵菌属Beauveria、单孢霉属Chloridium、外瓶霉属Exophiala、膝梗孢属Gonytrichum、白地霉属Geotrichum、粘束孢属Graphium、丝葚霉属Papulaspora、茎点霉属Phoma、毛癣菌属Trichophyton、毛束霉属Trichurus、沃德霉属Wardomyces、枝穗霉属Clonostachys各1个种。
在分离获得的各土壤真菌类群中,无孢类群真菌数量最多,占菌株总数的20.77%,是黑龙江省栽培药用植物根际土壤中最大的未知种类的真菌类群。Penicillium、Aspergillus、Trichoderma及Fusarium是黑龙江省栽培药用植物根际土壤中可明确真菌种类中数量最多的类群,分别占到菌株总数的16.93%、14.76%、8.46%和6.69%,是药用植物根际土壤中的优势种群。而Gliocladium、Chrysosporium、Mucor、Cladosporium、Acremonium、Rhizopus分别占菌株总数的5.02%、4.23%、4.13%、3.74%、2.36%和2.26%,是药用植物根际土壤中的亚优势种群。Paecilomyces和Alternaria是第三大优势种群,分别占1.48%和1.18%(表3)。Clonostachys、Exophiala、Geotrichum、Gonytrichum、Mortierella、Papulaspora、Trichophyton的菌株数仅为 1,是黑龙江省栽培药用植物根际土壤中的稀有真菌种群。
如表2所示,不同药用植物根际土壤真菌的种群组成和结构存在一定的差异,各真菌属种在不同的药用植物根际土壤中的优势度不同,不同的药用植物的根际土壤真菌区系结构也不同。在14种药用植物中,从五味子根际土壤中分离获得的真菌种类最多,达到33属真菌。无孢类群是14种药用植物根际土壤真菌中数量占优的类群,但它们最少属于5个菌落特征明显不同的类群。除无孢类群外,五味子根际土壤中的优势类群是青霉属、曲霉属和木霉属,它们占五味子根际土壤真菌总菌株数的相对分离频率分别为13.64%、11.36%和8.52%。从平贝母根际土壤中分离出26属真菌,其优势类群为青霉属、曲霉属和镰孢菌属,它们占平贝母根际土壤真菌总菌株数的相对分离频率分别为14.4%、14.4%和10.4%。从柴胡根际土壤中分离得到20属真菌,其优势类群为曲霉属、青霉属和木霉属,它们占柴胡根际土壤真菌总菌株数的相对分离频率分别为16.22%、12.16%和10.81%。其余11种药用植物根际土壤中除无孢类群外,青霉属、曲霉属、木霉属和镰孢菌属也为优势真菌类群。
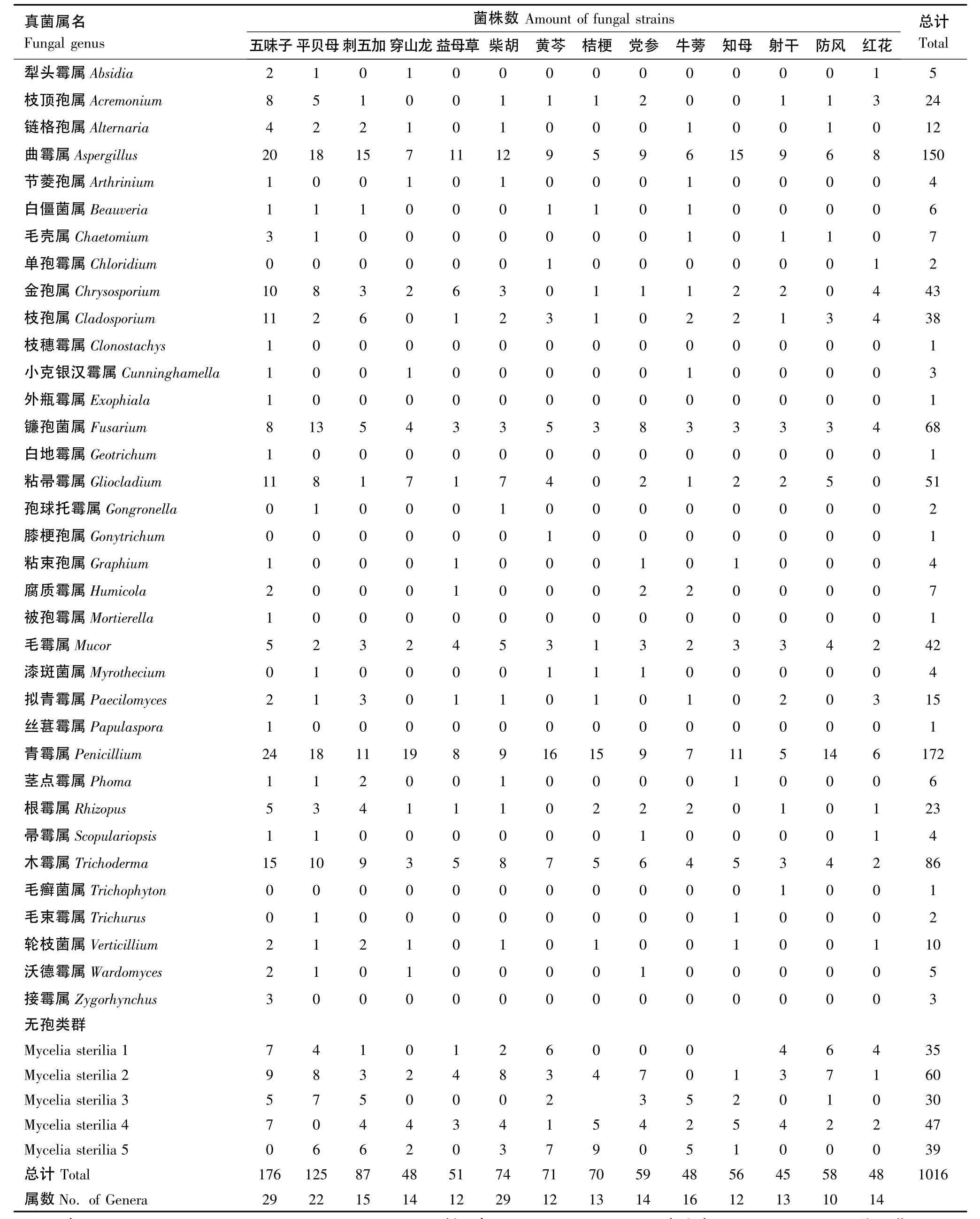
表2 黑龙江省药用植物根际土壤真菌种群组成Table 2 The fungal composition in rhizoshphere soil of medicinal plants in Heilongjiang Province
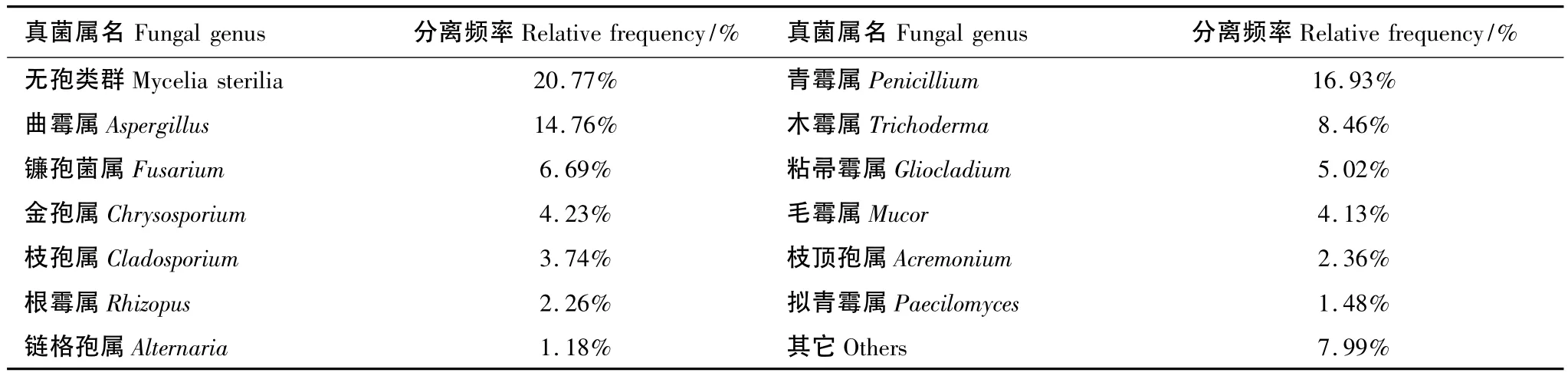
表3 黑龙江省药用植物根际土壤真菌优势种群的分离频率Table 3 The isolating frequencies of the major fungal genera in the rhizosphere soil of medicinal plants in Heilongjiang Province
2.2 黑龙江省3种主要药用植物根际土壤真菌种群结构组成与多样性分析
五味子、平贝母、柴胡是黑龙江省栽培区域最广泛和栽培面积最大的药用植物,它们在伊春、铁力、绥化、哈尔滨、牡丹江和佳木斯地区均有种植,且种植面积较大。以这3种药用植物为目标植物来分析它们的根际土壤真菌多样性。
2.2.1 不同地区3种主要药用植物根际土壤真菌多样性特点
在黑龙江省6个采样地点间的药用植物根际土壤真菌种群的多样性水平存在差异(表4)。从真菌属的数量上看,黑龙江省伊春地区的最高,共分离到25属真菌,而绥化的最低,仅分离到14属真菌。伊春地区真菌多样性指数(H')和丰富度指数(R)均最高,分别为2.9574和5.6683,而佳木斯地区真菌均匀度指数(J)最高,为 0.9200。

表4 黑龙江省6个采样地区3种主要药用植物根际土壤真菌种群的多样性指数Table 4 Diversity indices of fungal communities in rhizosphere soil of three main medicinal plants in six sampling regions of Heilongjiang Province
2.2.2 不同地区3种主要药用植物根际土壤真菌组成的相似性
在黑龙江省6个采样地点间药用植物根际土壤真菌的种群相似性系数在0.3704—0.6315之间,最高值为牡丹江与绥化的0.6315,最低值为牡丹江与哈尔滨的0.3704(表5)。根据Jaccard相似性原理,各地区之间的真菌相似性系数处于中等不相似与中等相似之间。总体来看,黑龙江省6个采样地区的药用植物根际土壤真菌种群相似性较低。

表5 黑龙江省6个采样地区3种主要药用植物根际土壤真菌相似性系数(Cj)Table 5 Similarity coefficients of the fungi isolated from rhizosphere soil of three main medicinal plants in six sampling regions of Heilongjiang Province
3 讨论
关于药用植物内生真菌多样性及微生物群落多样性的系统研究报道较多[17-19],而有关药用植物根际土壤真菌多样性的系统研究报道较少。本研究从黑龙江省6个地区栽培的药用植物根际土壤样品中分离和鉴定出真菌35属86种,对不同的药用植物以及不同地区药用植物根际土壤真菌的种类组成、区系分布和多样性特点进行了比较分析。研究结果表明,青霉属Penicillium、曲霉属Aspergillus、木霉属Trichoderma及镰孢菌属Fusarium真菌是黑龙江省药用植物根际土壤真菌的优势类群,这与已有的关于药用植物根际土壤真菌多样性的报道相似[20-21]。不同药用植物根际土壤中存在丰富的真菌资源,但其真菌种类组成、数量和分布具有较大的差异性。五味子根际土壤真菌中青霉属、曲霉属和木霉属为优势菌群,与潘争艳等[22]对辽宁栽培五味子根际土壤真菌的研究结果相似。不同种类药用植物根际土壤真菌的种类组成存在很大差异,且相同种类真菌的数量也存在较大差异。这种差异性可能是由不同药用植物根系分泌物的不同所造成的[23-24]。
多样性指数用来分析真菌种群或群落的多样性,丰富度指数用来反映真菌种属丰富性特点,均匀度指数是反映真菌种群分布特点的重要指标,而相似性系数是反映两个不同地区真菌种群组成相似程度的重要指数。本文以黑龙江省主产的五味子、平贝母和柴胡为典型代表进行分析,发现栽培地区不同,这3种药用植物的根际土壤真菌种群多样性水平的差异较大,相似程度较低。物种多样性指数、丰富度指数和均匀度指数与其生境中的土壤环境、植被以及人为因素(如栽培措施和管理水平)影响密切相关。伊春、铁力、绥化和牡丹江虽都地处山区,林地面积广,土壤肥沃,富含有机质,但是,伊春的五味子、平贝母和柴胡种植方式以林下种植为主,且历史悠久;绥化和牡丹江地区有农田种植和林下种植两种方式,而铁力地区却以农田种植为主,林下种植为辅,种植方式的不同导致其根际土壤真菌多样性水平存在差异,相似性程度较低。在哈尔滨地区这3种药用植物种植方式以农田种植方式为主,种植区土壤肥沃,水源充足,气候适宜,所以,其根际土壤真菌多样性水平较高。而在佳木斯地区它们的种植方式虽为农田种植,但种植区位于土壤相对贫瘠地带,水源也不充足,所以其根际土壤真菌多样性水平低,但是,其均匀度指数最高,说明该地区虽真菌种类资源不丰富,但分布却很均匀,这可能是由于种植区域内土壤生态环境差异不大造成的。虽然五味子、平贝母和柴胡在哈尔滨和佳木斯均为农田种植,但是,由于哈尔滨气候条件、种植区地理位置较佳木斯佳,其相似性程度较低。综上所述,药用植物的种植方式对其根际土壤真菌多样性水平存在较大影响,同时气候条件和土壤环境对其土壤真菌多样性水平也有一定的影响。本文试验结果反映了栽培生境不同对药用植物根际土壤真菌多样性存在不同程度的影响,与陈曦等[20]和杨皓[25]等的研究结论相一致。由于本研究采用传统的土壤真菌分离及形态学鉴定方法,可能导致不可培养或菌落生长较慢的真菌菌株被忽视。另外,本研究没有对不产孢类群真菌菌株进行确切鉴定(如采用DNA分子序列分析等),所以,导致本研究获得的黑龙江省药用植物根际土壤真菌多样性指数相对低些。
木霉菌Trichoderma和粘帚霉菌Gliocladium是非常重要的生防菌,由于它们具有生长迅速和具有拮抗和重寄生的优势特点,它们对土壤中的许多病原菌具有拮抗生防作用[26-27],已有较多报道利用木霉菌和粘帚霉菌防治植物土传病害,且取得较好防效[28-29]。本研究从黑龙江省药用植物根际土壤中分离得到木霉属8种86株和粘帚霉属2种51株,有望从中筛选到可用于植物土传病害防治的有效生防菌株。另外,这些年来关于利用真菌转化药用植物活性成分的研究报道很多,而且从中筛选出了具有较高生物活性的真菌菌株[30-32],这些类群的真菌通常能从药用植物的根际土壤中分离获得。不同药用植物的根系分泌物不同,其根际土壤中可能蕴藏着具有转化药用植物活性成分的功能真菌资源。本研究分离获得的大量真菌菌株也为今后开展微生物转化中药活性成分研究提供了丰富的真菌资源。
[1] Zhang W,Wei H L,Gao H W,Hu Y G.Advances of studies on soil microbial diversity and environmental impact factors.Chinese Journal of Ecology,2005,24(1):48-52.
[2] Denison R F,Bledsoe C,Kahn M,O'Gara F,Simms E L,Thomashow L S.Cooperation in the rhizosphere and the“free rider”problem.Ecology,2003,84(4):838-845.
[3] Barea J M,Pozo M J,Azcón R,Azcón-Aguilar C.Microbial co-operation in the rhizosphere.Journal of Experimental Botany,2005,56(417):1761-1778.
[4] Berg G,Smalla K.Plant species and soil type cooperatively shape the structure and function of microbial communities in the rhizosphere.FEMS Microbiology Ecology,2009,68(1):1-13.
[5] Raaijmakers J M,Paulitz T C,Steinberg C,Alabouvette C,Moënne-Loccoz Y.The rhizosphere:a playground and battlefield for soilborne pathogens and beneficial microorganisms.Plant and Soil,2009,321(1/2):341-361.
[6] Zhang Z Y,Lin W X.Continuous cropping obstacle and allelopathic autotoxicity of medicinal plants.Chinese Journal of Eco-Agriculture,2009,17(1):189-196.
[7] Ding W L.A Colored Atlas of Medicinal Plant Pests and Pest Control.Beijing:China Agriculture Press,2002:1-79.
[8] Gao W W,Jiao X L,Bi W,He C N.Variation of ginsenosides in infected roots of American ginseng(Panax quinquefolium).China Journal of Chinese Materia Medica,2008,33(24):2905-2907,2953.
[9] Ding W L,Cheng H Z,Chen J.Research on preventing the medicinal plant diseases withTrichoderma harzianumpreparation.China Journal of Chinese Materia Medica,2003,28(1):24-27.
[10] Zhang W,Sun X D,Zhang P,Lü G Z.Screening of the active fungal strains with high specificity to biotransform ginsenoside Rb1 into Rd.Mycosystema,2011,30(2):305-311.
[11] Domsch K H,Gams W,Anderson T H.Compendium of Soil Fungi.London:Academic Press,1980:1-859.
[12] Gams W,Bissett J.Morphology and identification ofTrichoderma//Kubicek C P,Harman G E,eds.TrichodermaandGliocladium.Volume 1.Basic Biology,Taxonomy and Genetics.London:Taylor and Francis Ltd,1998:3-34.
[13] Pei K Q.Taxonomic Studies on the Psychrophilic and Psychrotrophic Mucorales from China[D].Beijing:Institute of Microbiology,Chinese Academy of Sciences,1998:1-87.
[14] Qi Z T,Kong H Z,Sun Z M.Flora Fungorum Sinicorum Vol.5(Aspergilluset Teleomorphi Cognati).Beijing:Science Press,2003:1-173.
[15] Leslie J F,Summerell B A,Bullock S.TheFusariumLaboratory Manual.Oxford:Blackwell Publishing,2006:1-278.
[16] Kong H Z,Wang L.Flora Fungorum Sinicorum Vol.35(Penicilliumet Teleomorphi Cognati).Beijing:Science Press,2007:1-269.
[17] Huang W Y,Cai Y Z,Hyde K D,Corke H,Sun M.Biodiversity of endophytic fungi associated with 29 traditional Chinese medicinal plants.Fungal Diversity,2008,33:61-75.
[18] Sun J Q,Guo L D,Zang W,Ping W X,Chi D F.Diversity and ecological distribution of endophytic fungi associated with medicinal plants.Science in China Series C:Life Sciences,2008,38(5):475-484.
[19] Karthikeyan B,Jaleel C A,Lakshmanan G M A,Deiveekasundaram M.Studies on rhizosphere microbial diversity of some commercially important medicinal plants.Colloids and Surfaces B:Biointerfaces,2008,62(1):143-145.
[20] Chen X,Sun X D,Bi S Y,Zhang W,Yang H,Lü G Z,Xin X,Sun X Y.Fungal diversity in rhizosphere soil of medicinal plants in Liaoning Province.Mycosystema,2010,29(3):321-328.
[21] Zhang R,Liu Z H,Yang H,Bai X M,Xu H Y.Analysis of ecological characteristics and classification of fungal community in rhizosphere soil of ginseng in Jilin province.Journal of Jilin Agricultural Sciences,2008,33(1):47-50.
[22] Pan Z Y,Fu J F,Zhou R J,Wang H.Preliminary study of soil fungi inSchisandra chinensisgarden and screening antagonism strains.Journal of Jilin Agricultural University,2007,29(6):636-639.
[23] Bais H P,Weir T L,Perry L G,Gilroy S,Vivanco J M.The role of root exudates in rhizosphere interactions with plants and other organisms.Annual Review of Plant Biology,2006,57(1):233-266.
[24] Broeckling C D,Broz A K,Bergelson J,Manter D K,Vivanco J M.Root exudates regulate soil fungal community composition and diversity.Applied and Environmental Microbiology,2008,74(3):738-744.
[25] Yang H.Preliminary Study on the Rhizosphere Fungi of Some Medicinal Plants in Sichuan[D].Chengdu:Sichuan Agricultural University,2008:1-87.
[26] Papavizas G C.TrichodermaandGliocladium:biology,ecology,and potential for biocontrol.Annual Review of Phytopathology,1985,23(1):23-54.
[27] Whipps J M.Microbial interactions and biocontrol in the rhizosphere.Journal of Experimental Botany,2001,52(Suppl 1):487-511.
[28] Whipps J M,Davies K G.Success in biological control of plant pathogens and nematodes by microorganisms//Gurr G,Wratten S D,eds.Measures of Success in Biological Control.Dordrecht,the Netherlands:Kluwer Academic Publishers,2000:231-269.
[29] Whipps J M,Lumsden R D.Commercial use of fungi as plant disease biological control agents:status and prospects//Butt T M,Jackson C W,Magan N,eds.Fungi as Biocontrol Agents:Progress,Problems and Potential.Wallingford,UK:CABI Publishing,2001:9-22.
[30] Ning L L,Qu G Q,Ye M,Guo H Z,Bi K S,Guo D.Cytotoxic biotransformed products from triptonide byAspergillus niger.Planta Medica,2003,69(9):804-808.
[31] Ge W Z,Li N,Shan L H,Liu H M.Microbial transformation of 4-ene-3-one steroids byMucor racemosus.Acta Microbiologica Sinica,2007,47(3):540-543.
[32] Zhou W,Yan Q,Li J Y,Zhang X C,Zhou P.Biotransformation ofPanaxnotoginseng saponins into ginsenoside compound K production byPaecilomyces bainiersp.229.Journal of Applied Microbiology,2008,104(3):699-706.
参考文献:
[1] 张薇,魏海雷,高洪文,胡跃高.土壤微生物多样性及其环境影响因子研究进展.生态学杂志,2005,24(1):48-52.
[6] 张重义,林文雄.药用植物的化感自毒作用与连作障碍.中国生态农业学报,2009,17(1):189-196.
[7] 丁万隆.药用植物病虫害防治彩色图谱.北京:中国农业出版社,2002:1-79.
[8] 高微微,焦晓林,毕武,何春年.罹病西洋参根内主要人参皂苷含量的变化.中国中药杂志,2008,33(24):2905-2907,2953.
[9] 丁万隆,程惠珍,陈君.应用木霉制剂防治几种药用植物病害的研究.中国中药杂志,2003,28(1):24-27.
[10] 张薇,孙晓东,张萍,吕国忠.专一转化人参二醇类皂苷Rb1为Rd的真菌菌株的筛选.菌物学报,2011,30(2):305-311.
[13] 裴克全.中国低温毛霉的分类学研究[D].北京:中国科学院微生物研究所,1998:1-87.
[14] 齐祖同,孔华忠,孙增美.中国真菌志第五卷(曲霉属及相关有性型).北京:科学出版社,2003:1-173.
[16] 孔华忠,王龙.中国真菌志第三十五卷(青霉属及其相关有性型属).北京:科学出版社,2007,1-269.
[18] 孙剑秋,郭良栋,臧威,平文祥,迟德富.药用植物内生真菌多样性及生态分布.中国科学C辑:生命科学,2008,38(5):475-484.
[20] 陈曦,孙晓东,毕思远,张薇,杨红,吕国忠,辛鑫,孙晓燕.辽宁地区药用植物根际土壤真菌多样性的研究.菌物学报,2010,29(3):321-328.
[21] 张睿,刘志恒,杨红,白晓穆,徐怀友.吉林省人参根际土壤真菌群落生态特征及区系分析.吉林农业科学,2008,33(1):47-50.
[22] 潘争艳,傅俊范,周如军,王慧.五味子园根际真菌多样性初探及拮抗菌株筛选.吉林农业大学学报,2007,29(6):636-639.
[25] 杨皓.四川几种道地药材根际土壤真菌的初步研究[D].四川:四川农业大学,2008.
[31] 葛文中,李楠,单丽红,刘宏民.总状毛霉对4-烯-3-酮甾体的生物转化研究.微生物学报,2007,47(3):540-543.
猜你喜欢
中国现代中药(2022年3期)2022-04-11
中国土壤与肥料(2021年5期)2021-12-02
中国现代中药(2021年9期)2021-11-16
特产研究(2019年3期)2019-09-21
江苏农业科学(2019年5期)2019-09-02
天然产物研究与开发(2018年3期)2018-05-07
广东农业科学(2017年5期)2017-08-29
少儿科学周刊·儿童版(2017年5期)2017-06-29
中国蔬菜(2016年8期)2017-01-15
应用海洋学学报(2014年1期)2014-11-22
