
松嫩草地66种草本植物叶片性状特征
2013-12-19 07:30宋彦涛周道玮
生态学报 2013年1期
宋彦涛,周道玮 ,王 平,李 强
(1.中国科学院东北地理与农业生态研究所,长春 130102;2.东北师范大学草地科学研究所,长春 130024;3.东北师范大学城市与环境科学学院;长春 130024)
植物性状反映植物对生长环境的响应和适应,并应用于全球变化、古植被恢复和古气候定量重建、环境监测与评价、生态保护和恢复等研究[1]。而绿色叶片是陆地生态系统功能的基础[2],它们的形态和生理性状与陆地生态系统的结构和功能有着密切的联系[3]。不同叶片性状之间经常表现出相关关系,并且叶片性状之间的关系普遍存在于植物种群、群落和生物区系中,能够明显的反映出植物对气候和其它环境限制的趋同适应[4-5]。
松嫩草地位于东北腹地,四周环绕着大兴安岭、小兴安岭、长白山和松辽分水岭,是一个独立的自然区[6-7]。研究表明松嫩草地为土壤决定的植被类型[6],有独特的群落构成和植物区系,土壤中盐离子含量普遍高,植物相应地表现出抗盐耐盐特征[6-7],但松嫩草地草本植物叶片性状表现及其相关关系与其它草地的分异性和自身的趋同适应研究鲜有报道。本文以松嫩草地66种常见草本植物为研究对象,调查植物叶片的干物质含量(LDMC)、比叶面积(SLA)、叶片厚度(LT)、叶片氮含量(LNC)、叶片磷含量(LPC)、叶绿素含量(Chl)和类胡萝卜素含量(Cd)等7个叶片性状(表1),揭示该区草本植物叶片性状特征,分析性状间的相互关系,比较不同功能群之间性状的差异性,以期为区域研究和草地合理利用提供科学依据。

表1 植物性状缩写和单位Table 1 Definitions of abbreviations,acronyms,and units of plant traits
1 研究区概况与研究方法
1.1 研究区自然概况
研究地点位于中国科学院长岭草地农牧生态研究站(44°33'N,123°31'E,海拔145 m)。该区属于温带大陆性季风气候:春季干旱多风且降水稀少,地表蒸发剧烈;夏季降水集中,冬季降雪较少。年均温4.9℃,≥10℃积温2920℃,无霜期140—160 d,年日照时数2800 h。年降水量300—500 mm,集中在6—9月,这个时期降水量占总降水量的70%。年蒸发量1600.2 mm,约是降水量的3.5倍。该地区地势平坦,海拔140—160 m,以低地平原为主,有带状固定沙丘分布。地带性土壤为黑钙土,pH为7.5—9。
1.2 实验方法
2008年8月份,在中国科学院长岭草地农牧生态研究站附近的草甸共采集19科54属66种草本植物。划分为4个植物功能群[8]:(1)多年生根茎禾草(PR;n=9),(2)多年生丛生禾草(PB;n=5),(3)多年生杂类草(PF;n=39),(4)1年生或2年生草本(AB;n=13)。
每种植物采集新长出完全展开的且没有被啃食的成熟叶片(除去叶柄)约30 g带回实验室105℃恒温杀青18 min,75℃恒温烘干至恒重,粉碎,保存待测。按照相同标准每种植物采集10片叶片,装入有少量去离子水的封口袋中,放入装有冰的保温箱中带回实验室,黑暗条件下保存于4℃恒温冰箱中24 h。取出后用滤纸迅速擦干叶片表面水分,万分之一天平称量叶片的饱和鲜重;测量叶片厚度(LT);照相,用SigmaScan Pro 4求算每个叶片的叶面积;最后把叶片放入75℃烘箱烘干24 h后称重。SLA=叶片面积/叶片干重,LDMC=叶片干重/叶片饱和水鲜重。
LNC用凯式定氮法测定,LPC用NaOH熔融-钼锑钪比色法测定[9]。叶绿素a、b,Cd用分光光度法测定,Chl=叶绿素 a+叶绿素 b[10]。
1.3 数据处理
叶片性状的相关性用Pearson相关分析(双尾检验)和多元回归分析,不同功能群间叶片性状的差异用单因素方差分析(方差不齐时进行数据转换)和LSD多重比较法检验均值间的差异,显著性水平a=0.05,所有统计使用SPSS软件 (2004,ver.13.0;SPSS Inc.,USA)完成。
2 结果
2.1 叶片性状特征
66种植物叶片性状值变异较大,最大值是最小值的3.5倍以上,变异系数超过30%(表2)。其中,叶片厚度的变异最大,最大值是最小值的9.5倍,变异系数接近60%;其次是叶片磷含量(CV=36.5%)和叶片干物质含量(CV=35.4%);比叶面积、叶片氮含量、叶绿素含量和类胡萝卜素含量的变异系数在31%—33.5%之间。
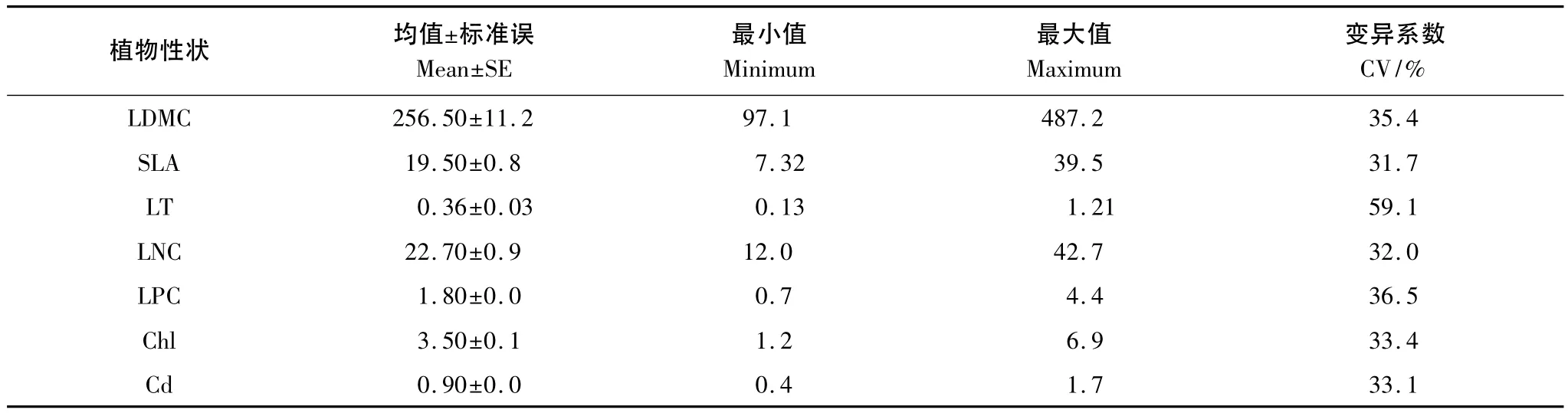
表2 松嫩草地66种草本植物叶片性状特征(n=66)Table 2 Leaf traits characteristic of 66 herbaceous plants in Songnen grassland
2.2 叶片性状之间的相关关系
66种植物的叶片干物质含量与叶片磷含量没有显著的相关性(P=0.070),与其它植物性状都呈现显著的负相关关系;比叶面积和叶片厚度呈现显著的负相关(P=0.008),与叶片氮含量、叶片磷含量、叶绿素和类胡萝卜素呈现显著的正相关;叶片厚度与叶片氮含量、叶片磷含量、叶绿素和类胡萝卜素不相关;叶片氮含量与叶片磷含量、叶绿素和类胡萝卜素呈现显著的正相关;叶片磷含量与叶绿素和类胡萝卜素呈现显著的正相关;叶绿素与类胡萝卜素呈现显著的正相关(表3)。
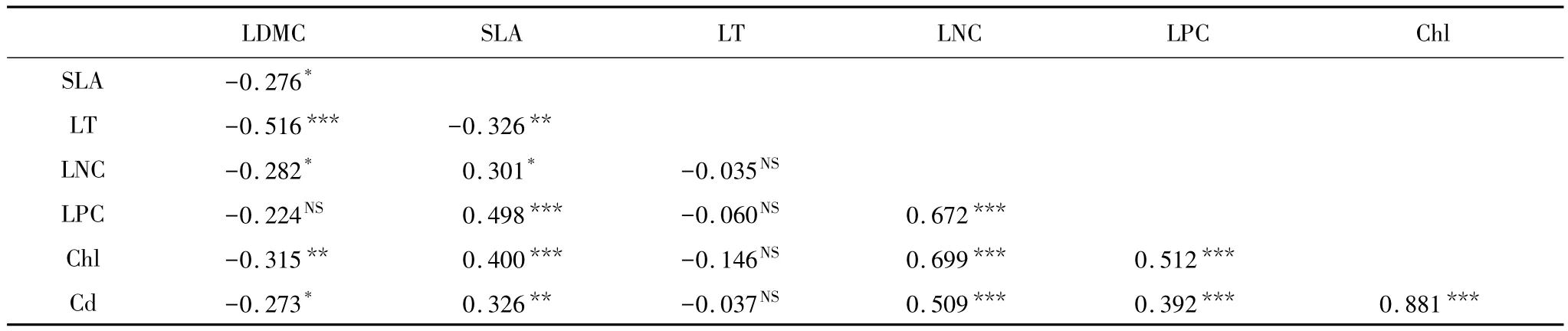
表3 叶片性状的相关性(n=66)Table 3 Correlation coefficient among leaf traits
多元回归显示(图1),叶片厚度与叶片干物质含量和比叶面积都有显著的负相关关系 (LT=-0.002LDMC-0.017SLA+1.092),即对任意的叶片干物质含量,叶片厚度随比叶面积的增加而降低;对任意的比叶面积,叶片厚度随叶叶片干物质含量的增加而降低。绿素含量与叶片氮含量和比叶面积有显著的正相关关系(Chl=0.101LNC+0.039SLA+0.406),即对任意的叶片氮含量,叶绿素随比叶面积的增加而增加;对任意的比叶面积,叶绿素随叶片氮含量的增加而增加。
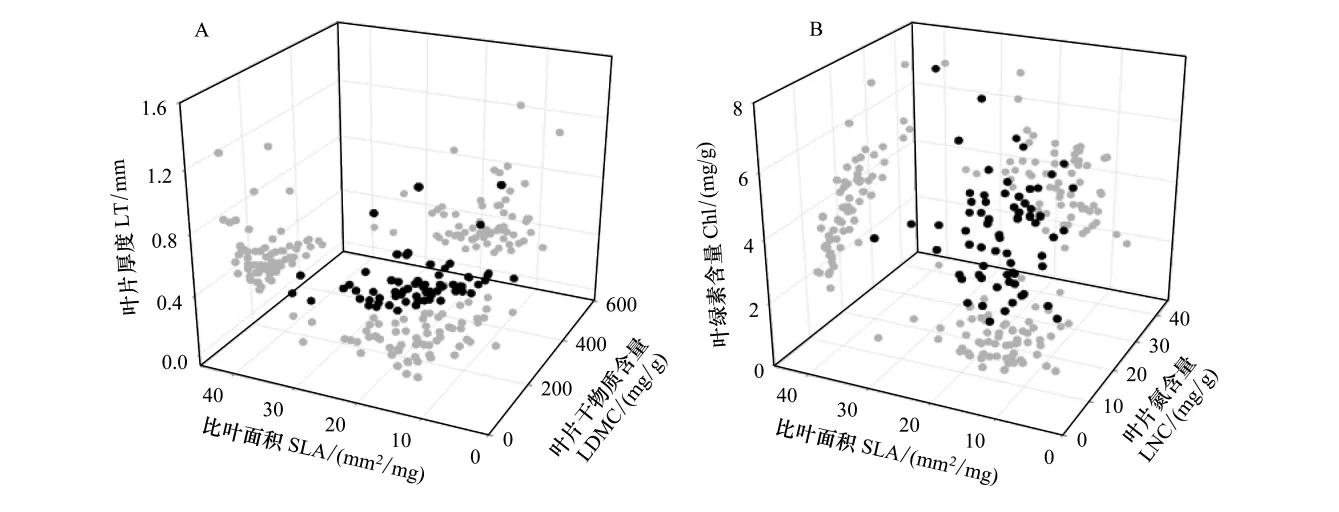
图1 叶片性状关系3维图Fig.1 Leaf trait relationships in three-dimensional space
2.3 不同功能群植物叶片性状的差异
方差分析结果表明,叶绿素和类胡萝卜素在不同功能群间没有显著差异(P>0.05),其它性状在不同功能群间差异显著。多年生根茎禾草和多年生丛生禾草的叶片干物质含量显著的高于多年生杂类草和1年生或2年生草本;1年生或2年生草本叶片的比叶面积显著的高于其它3个功能群;多年生杂类草的叶片厚度显著的高于多年生根茎禾草,多年生丛生禾草和1年生或2年生植的叶片厚度与其它两个功能群差异不显著;多年生杂类草和1年生或2年生草本叶片的氮含量显著的高于多年生根茎禾草,与多年生丛生禾草差异不显著;1年生或2年生草本叶片的磷含量显著高于其它3个功能群(图2)。
3 讨论
3.1 松嫩草地草本植物叶片性状特征
植物性状反映了植物对气候和土壤等环境因子适应的重要信息[1],如叶片干物质含量和比叶面积与植物获取资源的能力密切相关[11-12]。松嫩草地植物与邻近的科尔沁沙地植物叶片性状相比,其比叶面积高于科尔沁沙地植物,而叶片干物质含量和叶片厚度低[13-15]。因为松嫩草地属于温带半湿润季风气候区,且雨热同季[7],而科尔沁沙地属于温带半干旱气候类型(年均温6.5℃,年均降水量360 mm,年蒸发量1935 mm)[15],相对干旱贫瘠,可利用资源少。比叶面积高的植物适应资源丰富的环境;相反,比叶面积低的植物适应了降水少、蒸发量大、土壤贫瘠的环境[15]。同样,松嫩草地植物叶片相对低的干物质含量和叶片厚度也是适应水热和土壤条件相对较好的结果。松嫩草地草本植物叶片厚度(变异系数59.1%)比其它叶片性状变异大(表2),可能因为不同植物间叶片形态差异很大及同种内也存在个体变异,同时叶片厚度值相对较小,导致叶片厚度测量误差较大[12,14]。

图2 不同植物功能群叶片性状比较Fig.2 Comparison of leaf traits among different plant functional groups
松嫩草地草本植物叶片氮磷含量分别为22.7 mg/g和1.8 mg/g(表2)。与其它地区相比,松嫩草地植物叶片的氮含量低于内蒙古草地、新疆草地和西藏草地,与科尔沁沙地植物叶片氮含量相当,高于全国陆生植物;叶片磷含量低于科尔沁沙地,高于内蒙古草地、西藏草地和全国陆生植物,与新疆草地相当;氮磷比除了高于科尔沁沙地外,均低于其它地区和全国陆生植物[15-18](表4)。与中国陆生植物和草原区植物叶片氮磷含量对比,松嫩草地植物叶片氮磷比低主要归因于低的叶片氮含量和高的叶片磷含量。氮和磷元素是植物生长发育最主要的限制因子,两者之间存在着重要的相互作用[4,17-18]。植物氮磷含量取决于土壤养分供应和植物氮磷需求间的动态平衡,植物氮磷比可以判断植物生长受限状况[19-20]。欧洲40个地点施肥实验表明,氮磷比大于16表示植被生长受磷限制,氮磷比小于14表示受氮限制,氮磷比在14—16时,受氮磷的共同限制[20],Gusewell[21]通过对大量研究结果综述认为氮磷比>20和<10作为植被水平氮磷限制的评价标准。松嫩草地草本植物叶片氮磷比为13.0,对比中国其它草原区植物氮磷含量和氮磷比(表4),综合判断松嫩草地相对于中国其它草原区来说磷相对丰富,氮相对缺乏。

表4 松嫩草地草本植物叶片氮磷质量浓度和氮磷比与其它研究结果比较Table 4 Comparisons between leaf N,P and N∶P of Songnen grassland and other studies on mass basis
3.2 松嫩草地草本植物叶片性状之间的相关关系
量化植物功能性状之间的关系,特别是多个物种和不同生境的植物叶片性状的关系是植物功能生态学研究要的重要内容之一[1,22]。研究表明,叶片性状之间相互关联,如比叶面积、光合能力、暗呼吸速率、叶片氮磷含量等相互之间都有正相关关系[2,23-25]。在松嫩草地,分析发现比叶面积、叶片干物质含量、叶片氮含量之间都有显著的相关关系(表3,图1)。叶片干物质含量反映的是叶片的组织密度,快生长植物的组织密度低,干物质含量低,比叶面积高[11],与其它性状间通常是负相关关系。
叶片厚度与叶片干物质含量和比叶面积都有极显著的负相关关系(表3),Vile等[12]通过对11个地点1039对数据的验证指出用比叶面积和叶片干物质含量乘积的倒数((SLA×LDMC)-1)能够很好的估算叶片厚度。结果显示用叶片厚度做因变量,叶片干物质含量和比叶面积做自变量,建立多元线性回归方程,也能很好的预测叶片厚度(图1)。但是该方程在其它地区是否适用,需要相应的数据数据进一步验证。
叶绿素和类胡萝卜素是主要的光合色素,能够指示植物叶片的光合能力,类胡萝卜素还能使激发态的叶绿素失活,避免多余的吸收能量对叶绿素的伤害[26-27]。与其它研究结果一致,叶绿素与叶片的氮含量有显著的正相关关系[28],且叶绿素和类胡萝卜素与除叶片厚度之外的其它叶片性状都有显著的相关关系。在叶片性状之间的相关系数中,叶绿素和类胡萝卜素的相关系数最大(r=0.881,表3),且叶绿素可以通过SPAD-502(日本,大阪)容易、快速的测量[29],因而可以用叶绿素来替代类胡萝卜素。
3.3 松嫩草地草本植物功能群叶片性状的差异
植物功能群是对环境有类似响应和对生态系统功能有相似作用的不同物种组合[30],不同功能群植物对环境的响应或生态系统功能存在差异[31]。在内蒙古典型草原区,根据植物的生活型,划分为多年生根茎禾草、多年生丛生禾草、多年生杂类草、灌木或半灌木和1年生或2年生植物5个功能群[8]。然而,松嫩草地的结果表明,多年生根茎禾草和多年生丛生禾草的7个叶片性状均差异不显著(图2)。调查的多年生根茎禾草多属C4植物,多年生丛生禾草多属C3植物。一般来说,在相对高温、干旱和高光条件下,C4植物比C3植物有高的羧化作用和资源利用率[3,32]。然而,松嫩草地属于C3、C4植物分布的交错区,地处温带,仅在7—8月份能满足C4植物生理活动对强光和高温的需求,而且该区在7—8月份降雨最多[7]。在这样的条件下,C4植物的优势很难表现出来。这可能是C3、C4多年生禾草的叶片性状间也没有显著的差异的原因。由此,松嫩草地多年生禾草划分一个功能群可能更合理。
叶片干物质含量和比叶面积是植物功能生态学研究中最常用的性状,且叶片干物质含量和比叶面积的倒数呈正比关系[1,22]。它们在植物叶片功能中发挥着重要作用,能反映植物对资源获取和利用情况[11],研究指出叶片干物质含量高,比叶面积小的植物资源获取能力强,更能在逆境中占优势[33]。松嫩草地多年生禾草(多年生根茎禾草和多年生丛生禾草)植物叶片干物质含量高于多年生杂类草和1年生或2年生草本,比叶面积低于多年生杂类草和1年生或2年生草本(图2),表明多年生禾草比其它功能群植物更能适应该区环境。在一定程度上从植物叶片功能生态角度解释了松嫩草地多年生禾草占优势的原因。1年生或2年生草本叶片寿命短、生长速率快、高生殖投入,所以该功能群叶片干物质含量低,比叶面积和叶片厚度大;1年生或2年生草本需要更多的磷素用于高比例的繁殖分配,导致叶片氮磷含量比叶片寿命长、生长慢的功能群高[18,21]。尤其是叶片磷含量,显著高于其它功能群(图2)。
松嫩草地草本植物叶绿素和类胡萝卜素含量在各个功能群之间差异不显著(图2)。Zheng等[3]对黄土高原7个地点研究显示,该区乔木、灌木和草本3种生活型植物叶片的叶绿素含量有显著差异,而我们的研究区域内,温度、光照、降水等环境因子较为一致。草本植物不同功能群叶片叶绿素和类胡萝卜素对环境的响应相对不敏感,可能是不同植物功能群之间差异不显著的原因。需要进一步扩大研究区域和增加物种数量来比较不同功能群间的差异性。
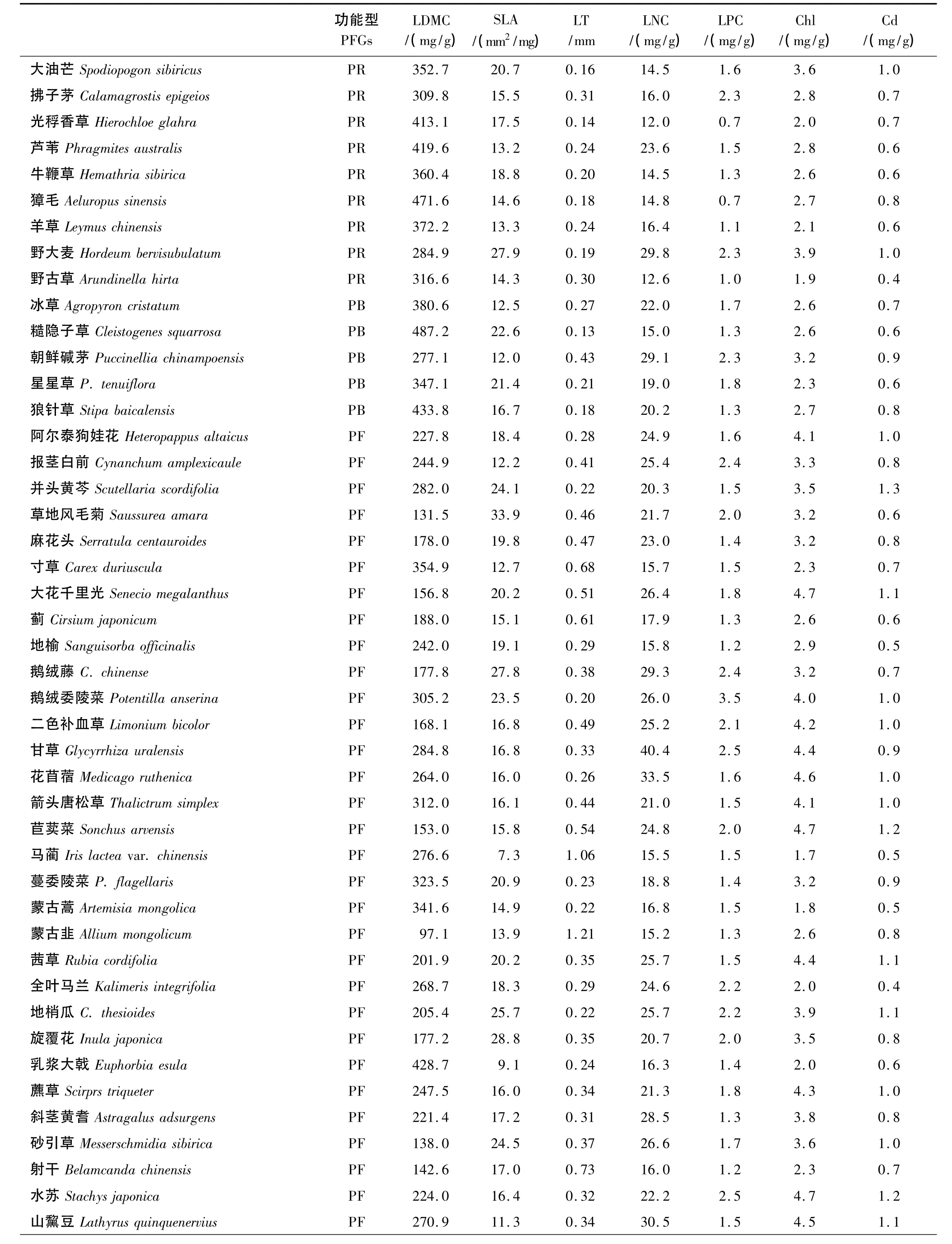
附表1 松嫩草地66种草本植物叶片性状Appendix table 1 Leaf traits of 66 herbage plant species in Songnen grassland,China

续表
[1] Meng T T,Ni J,Wang G H.Plant functional traits,environments and ecosystem functioning.Journal of Plant Ecology,2007,31(1):150-165.
[2] Wright I J,Reich P B,Westoby M,Ackerly D D,Baruch Z,Bongers F,Cavender-Bares J,Chapin T,Cornelissen J H C,Diemer M,Flexas J,Garnier E,Groom P K,Gulias J,Hikosaka K,Lamont B B,Lee T,Lee W,Lusk C,Midgley J J,Navas M L,Niinemets Ü,Oleksyn J,Osada N,Poorter H,Poot P,Prior L,Pyankov V I,Roumet C,Thomas S C,Tjoelker M G,Veneklaas E J,Villar R.The worldwide leaf economics spectrum.Nature,2004,428(6985):821-827.
[3] Zheng S X,Shangguan Z P.Spatial patterns of photosynthetic characteristics and leaf physical traits of plants in the Loess Plateau of China.Plant Ecology,2007,191(2):279-293.
[4] Reich P B,Oleksyn J.Global patterns of plant leaf N and P in relation to temperature and latitude.Proceedings of the National Academy of Sciences of the United States of America,2004,101(30):11001-11006.
[5] Wright I J,Reich P B,Cornelissen J H C,Falster D S,Groom P K,Hikosaka K,Lee W,Lusk C H,Niinemets Ü,Oleksyn J,Osada N,Poorter H,Warton D I,Westoby M.Modulation of leaf economic traits and trait relationships by climate.Global Ecology and Biogeography,2005,14(5):411-421.
[6] Zhou D W,Zhang Z X,Jin Y H,Wang P,Wang X Z.Regionalization and distribution pattern of vegetation of Northeast China.Chinese Journal of Plant Ecology,2010,34(12):1359-1368.
[7] Zheng H Y,Li J D.The Grassland Vegetation and Its Utilization and Conservation on the Songnen Plain.Beijing:Science Press,1993.
[8] Bai Y F,Han X G,Wu J G,Chen Z Z,Li L H.Ecosystem stability and compensatory effects in the Inner Mongolia grassland.Nature,2004,431(7005):181-184.
[9] Bao S D.Analysis of Soil Characteristics.3rd edn.Beijing:Chinese Agricultural Press,2005.
[10] Wang Y D,Liu N.Experimental Guidance for Plant Biology.Beijing:Higher Education Press,2001.
[11] Cornelissen J H C,Lavorel S,Garnier E,Díaz S,Buchmann N,Gurvich D E,Reich P B,ter Steege H,Morgan H D,van der Heijden M G A,Pausas J G,Poorter H.A handbook of protocols for standardised and easy measurement of plant functional traits worldwide.Australian Journal of Botany,2003,51(4):335-380.
[12] Vile D,Garnier É,Shipley B,Laurent G,Navas M L,Roumet C,Lavorel S,Díaz S,Hodgson J G,Lloret F,Midgley G F,Poorter H,Rutherford M C,Wilson P J,Wright I J.Specific leaf area and dry matter content estimate thickness in laminar leaves.Annals of Botany,2005,96(6):1129-1136.
[13] Li X L,Li X H,Jiang D M,Liu Z M,Wang H M,Ji L Z.Leaf morphological characters of 22 compositae herbaceous species in Horqin sandy land.Chinese Journal of Ecology,2005,24(12):1397-1401.
[14] Liu J H,Zeng D H,Lee D K.Leaf traits and their interrelationships of main plant species in southeast Horqin sandy land.Chinese Journal of Ecology,2006,25(8):921-925.
[15] Zhao H Y,Li Y L,Wang X Y,Mao W,Zhao X Y,Zhang T H.Variations in leaf traits of 52 plants in Horqin sand land.Journal of Desert Research,2010,30(6):1292-1298.
[16] He J S,Fang J Y,Wang Z H,Guo D L,Flynn D F B,Geng Z.Stoichiometry and large-scale patterns of leaf carbon and nitrogen in the grassland biomes of China.Oecologia,2006,149(1):115-122.
[17] He J S,Wang L,Flynn D F B,Wang X P,Ma W H,Fang J Y.Leaf nitrogen:phosphorus stoichiometry across Chinese grassland biomes.Oecologia,2008,155(2):301-310.
[18] Han W X,Fang J Y,Guo D L,Zhang Y.Leaf nitrogen and phosphorus stoichiometry across 753 terrestrial plant species in China.New Phytologist,2005,168(2):377-385.
[19] Zeng D H,Chen G S.Ecological stoichiometry:a science to explore the complexity of living systems.Chinese Journal of Plant Ecology,2005,29(6):1007-1019.
[20] Koerselman W,Meuleman A F M.The vegetation N ∶P ratio:a new tool to detect the nature of nutrient limitation.Journal of Applied Ecology,1996,33(6):1441-1450.
[21] Güsewell S.N:P ratios in terrestrial plants:variation and functional significance.New Phytologist,2004,164(2):243-266.
[22] Zhou D W.A phylogenic approach to comparative functional plant ecology.Acta Ecologica Sinica,2009,29(10):5644-5655.
[23] Wright I J,Groom P K,Lamont B B,Poot P,Prior L D,Reich P B,Schulze E D,Veneklaas E J,Westoby M.Leaf trait relationships in Australian plant species.Functional Plant Biology,2004,31(5):551-558.
[24] Wright I J,Reich P B,Cornelissen J H C,Falster D S,Garnier E,Hikosaka K,Lamont B B,Lee W,Oleksyn J,Osada N,Poorter H,Villar R,Warton D I,Westoby M.Assessing the generality of global leaf trait relationships.New Phytologist,2005,166(2):485-496.
[25] Reich P B,Walters M B,Ellsworth D S.From tropics to tundra:global convergence in plant functioning.Proceedings of the National Academy of Sciences of the United States of America,1997,94(25):13730-13734.
[26] Lewandowska M,Jarvis P G.Changes in chlorophyll and carotenoid content,specific leaf area and dry weight fraction in Sitka spruce,in response to shading and season.New Phytologist,1977,79(2):247-256.
[27] Veres S,Tóth V R,Láposi R,Oláh V,Lakatos G,Mészáros I.Carotenoid composition and photochemical activity of four sandy grassland species.Photosynthetica,2006,44(2):255-261.
[28] Gáaborcˇícik N.Relationship between contents of chlorophyll(a+b)(SPAD values)and nitrogen of some temperate grasses.Photosynthetica,2003,41(2):285-287.
[29] Bakker M A,Carreño-Rocabado G,Poorter L.Leaf economics traits predict litter decomposition of tropical plants and differ among land use types.Functional Ecology,2011,25(3):473-483.
[30] Gitay H,Nobel I R.What are functional types and how should we seek them? ∥Smith T M,Shugart H H,Woodward F I,eds.Plant Functional Types:Their Rrelevance to Ecosystem Properties and Global Change.Cambridge:Cambridge University Press,1997:3-19.
[31] Cabído M,Cabido M.Vive la différence:plant functional diversity matters to ecosystem processes.Trends in Ecology and Evolution,2001,16(11):646-655.
[32] Sage R F,Monson R K.The biogeography of C4photosynthesis:patterns and controlling factors∥Sage R F,Monson R K,eds.C4Plant Biology.San Diego:Academic Press,1999:313-373.
[33] Wilson P J,Thompson K,Hodgson J G.Specific leaf area and leaf dry matter content as alternative predictors of plant strategies.New Phytologist,1999,143(1):155-162.
参考文献:
[1] 孟婷婷,倪健,王国宏.植物功能性状与环境和生态系统功能.植物生态学报,2007,31(1):150-165.
[6] 周道玮,张正祥,靳英华,王平,王学志.东北植被区划及其分布格局.植物生态学报,2010,34(12):1359-1368.
[7] 郑慧莹,李建东.松嫩平原的草地植被及其利用保护.北京:科学出版社,1993.
[9] 鲍士旦.土壤农化分析(第三版).北京:中国农业出版社,2005.
[10] 王英典,刘宁.植物生物学实验指导.北京:高等教育出版社,2001.
[13] 李晓兰,李雪华,蒋德明,刘志民,王红梅,姬兰柱.科尔沁沙地22种菊科草本植物叶片形态特征研究.生态学杂志,2005,24(12):1397-1401.
[14] 刘金环,曾德慧,Lee D K.科尔沁沙地东南部地区主要植物叶片性状及其相互关系.生态学杂志,2006,25(8):921-925.
[15] 赵红洋,李玉霖,王新源,毛伟,赵学勇,张铜会.科尔沁沙地52种植物叶片性状变异特征研究.中国沙漠,2010,30(6):1292-1298.
[19] 曾德慧,陈广生.生态化学计量学:复杂生命系统奥秘的探索.植物生态学报,2005,29(6):1007-1019.
[22] 周道玮.植物功能生态学研究进展.生态学报,2009,29(10):5644-5655.
猜你喜欢
今日农业(2022年14期)2022-11-10
草地学报(2021年7期)2021-08-03
湖南农业大学学报(自然科学版)(2021年3期)2021-07-02
新疆农业科学(2020年1期)2020-02-14
南方农业·下旬(2017年11期)2018-01-09
中国饲料(2017年15期)2017-08-22
杂草学报(2015年2期)2016-01-04
中国麻业科学(2015年5期)2015-12-28
湖南农业科学(2014年14期)2014-02-27
环境工程技术学报(2011年4期)2011-09-05
