
河静黑叶猴果实性食物组成、选择及其对种子的扩散作用
2013-12-19 07:30阮海河鲁长虎
生态学报 2013年1期
阮海河,白 冰,李 宁,潘 扬,鲁长虎,*
(1.南京林业大学,南京 210037;2.越南林业大学,河内 100803)
叶猴以叶为主要食物,但在它们的食物资源中,果实是仅次于树叶的第二大食物资源[1,2],分布在马来西亚的印尼叶猴(Presbytis femoralis)取食的果实和种子种类达到其食谱的46%[3]。受所在生态环境的影响,叶猴取食果实的种类甚至高于树叶,如橡胶园和果园的托式叶猴(Presbytis thomasi),取食的果实和种子占58%,树叶仅占32%[4]。更多的叶猴食性研究发现,由于食物资源的时空变化,在嫩叶较少的季节,它们总会寻找一些固定的果实种类作为重要的补充食物[2,5]。
灵长类通过取食果实传播种子,因此对森林生态系动力学机制具重要作用[6-7],其对果实的选择受到果实特征,如大小[6]、色泽[7]、易处理程度[8]等影响,由此传播的种子与对植物更新的功能亦不相同。和其他灵长类动物一样,叶猴对果实的取食同样使它们成为种子的潜在传播者。在相关的种子传播研究中,大多关注于食果实灵长类,如长臂猿[9]、蜘蛛猴[10]和狐猴[11]等,而少有叶猴对种子扩散作用的报道。喀斯特地貌悬崖峭壁多,缺少地表径流,山体小且隔离程度高,与其他类型的森林相比,喀斯特森林的生态环境更为脆弱[12-13]。由于抗干扰和恢复能力低,喀斯特森林中种子扩散对维持植物种群的续存与更新具极其重要的作用。生活在其中的动物通过取食植物果实、传播种子,对稳定植物种群,促进更新与扩散起重要作用。迄今我们在动物对喀斯特森林的更新和扩散作用方面的了解还远远不够。
河静黑叶猴(Trachypithecus francoisi hatinhensis)是生活在喀斯特环境的灵长类动物,属越南特有种。已有研究者对其亲缘种,同样生活在喀斯特环境的黑叶猴(Trachypithecus francoisi)食性进行了详细的研究,其中涉及到果实类食物的选择及季节变化等[14-16]。迄今尚未出现叶猴类动物关于种子传播的研究报告。基于此,选择分布于越南风芽-格邦国家自然公园的河静黑叶猴,通过调查它们对果实的选择与取食,以及粪便中种子及传播距离,以期了解(1)河静黑叶猴的一年中果实类食物组成;(2)河静黑叶猴对果实种类及特征的选择偏好;(3)河静黑叶猴对哪些果实植物具潜在的种子传播作用。
1 材料与方法
1.1 研究区概况
风芽-格邦国家自然公园(17°21'12″—17°39'44″N ,105°57'53″—106°24'19″E)位于越南中部,面积 85000 hm2,西面与老挝接壤,为安南山脉的一部分,海拔50—1000 m。公园为喀斯特地貌的石山环境,植被以热带季雨林、亚热带常绿阔叶林及石山灌丛为主。石山丛相对海拔200—300 m,最高1200 m。风芽-格邦国家自然公园属湿热的热带气候特征,具有明显的旱雨季划分[17]。
1.2 研究方法
1.2.1 猴群跟踪
作为河静黑叶猴的主要分布区,风芽-格邦国家自然公园确定有16个河静黑叶猴群[18]。选择一个由18只个体组成的猴群为观察对象。该猴群由6只成年雄猴、9只成年雌猴及3只幼猴组成。
于2009年7月至2010年12月对目标猴群进行跟踪观察。观察开始于最初发现猴群的时刻。如果能确定前一天猴群的夜宿地,观察开始于猴群离开夜宿地,一直持续到猴群进入夜宿地。野外共跟踪观察标猴群269 d,每月不少于16 d,其中全天候跟踪观察不少于5 d。
1.2.2 果实食物及取食行为
跟踪过程中,用30×50的双筒望远镜在20—100 m距离对目标猴群进行观察。采用瞬时扫描法[19]对猴群的取食频次进行取样,扫描时间5 min,取样间隔10 min。为减小个体识别和移动带来的取样误差,取样时采取两种方法收集取食频次数据:(1)每天仅针对能辨别的个体进行取样,如第一天仅将成年雄猴作为取样个体,第二天仅将雌猴作为取样个体(数量2—4只不等)。确定目标个体后,(2)取样中观察人员分为两组,一组负责取样及记录,另一组则持续观察猴群的取食活动,在下一次取样时提供个体位置变化的信息。取样时记录每个取样个体采食的果实种类、频次并标记取食树。对于野外不能确定种类又无法接近的果树,则在跟踪结束后尽可能地收集猴群取食时掉落的果实和折断的枝叶,并用长焦镜头拍下果树照片带回鉴定。另外,通过收集粪便,鉴定粪便内的种子以补充取食的果实种类。以河静黑叶猴对该果实的取食频次与总的取食频次之比来表示其取食强度。
采用样带和样方法调查果实的可获得性。和其他植物物候调查不同,果实具有时效性的特征。除了季节间果实种类的差异,野外调查发现,并非每棵果实树都能挂果,且挂果时间亦有差异。为了尽可能多的统计到猴群活动区内结果的果实植物数量,每个月末,沿目标猴群的觅食路线设计一条约5 km,宽2 m的样带。设置样带时尽可能地包括当月目标猴群的觅食路线及取食树。另外,在同一区域内沿坡面设置10个50 m×6 m的样方,标记样带和样方内所有结果的果实树棵数。到野外调查后期,只标记新发现的没有标记的果实树。果实的可获得性指数(FAI)[14],计算方法为:FAI=Ni/N。其中,Ni为i种果树的棵数,N为标记的果实树总棵数。
1.2.3 果实特征
参照文献[20]的方法,将取食的果实划分为浆果、核果、裂果(成熟后果皮裂开的干果)、闭果(成熟后果皮不裂开的干果)和假种皮(裸子植物)5种类型;记录果实颜色(根据观察猴群取食时的果实颜色,将果实分为黄色、红色、绿色、褐色、白色、紫色、黑色与双色)和每颗果实包含的种子数(1—2、3—4、5—6、7—8、>8)。另外,每种果实随机取 50 颗,测量果实平均重量(<5 g、5—50 g、>50 g)和种子平均重量(≤0.1 g、0.1—1 g、1—3 g、3—5 g、≥5 g)。果实的处理难易程度划分为5类,即r1:手指即能压碎;r2:指甲能轻易剥开;r3:指甲很困难才能剥开;r4:用刀能轻松剥开;r5:用刀很困难才能剥开。
1.2.4 对种子的扩散作用
采用直接观察法和粪便法相结合的方式调查河静黑叶猴取食果实时对种子的处理方式。当对猴群进行扫描取样时,调查组另一名成员选择观察角度最好的一个个体,仔细观察目标猴在取食时是否吐出种子。对于无法观察到的种子处理过程,则通过粪便分析来确定,如粪便中发现有完好种子则认为该种子被整吞且可通过肠道扩散,如发现种子碎片则认为该种子被咀嚼。在条件允许的情况下,接近取食树在附近仔细寻找是否有丢弃的新鲜种子。于2009年7至12月和2010年5至12月期间野外收集目标猴群的粪便。沿猴群的觅食路径和行径方向寻找粪便并标记所在地点。跟踪过程中,如发现有个体排便,待猴群离开后在其栖位仔细寻找并标记。每天跟踪结束后,按当天跟踪路线返回,在返回过程中清除所有看到的粪便且只收集新鲜的粪便以尽可能地确保所收集的粪便为目标猴群的。采用水洗法辨别粪便中的种子并取样鉴定。由于1月至4月为旱冬晚期和旱春季节,野外果实稀少,没有进行粪便统计。
为了解猴群对种子的扩散作用,通过用激光测距仪测量有完整种子的粪便距相应树种的距离来评估猴群对某种果实的扩散距离。该方法虽然不能确定粪便中种子来自哪棵果树,但至少能够反映出种子能够被携带的最短距离。
1.2.5 数据分析
采用Mann&W hitney U test来比较取食果实的季节差异[14]。采用选择指数S分析黑叶猴对果实的选择偏好[21]:

其中,FCsp为果实的取食强度。即河静黑叶猴对该果实的取食频次/取食总频次;FAsp为果实的资源量。由于野外地形所限,不可能准确评估每种果实的资源量。所以,采用果实的可获得性(FAI)替代其资源量,这种方法尽管不能精确地提供环境中食物的丰富度,但可以反映出食物可获得性的估计值[14]。式中,当S>0.3(FCsp为FAsp的两倍)时,对该果实为高度选择;当S在-0.3和0.3之间(FCsp与FAsp值相近),对该种果实为中度选择:当S<-0.3(FCsp小于FAsp的一半)时,对该果实为低度选择;当FCsp=0时,S=-1,该种果实不被采食。
为了解河静黑叶猴对果实的选择偏好是否受到果实的可获得性的影响,采用Pearson相关性指数验果实的可获得性(FAI)与选择指数S间的关系。相关性水平设定为0.05。所有数据分析均在Microsoft Excel和SPSS 13.0统计软件上完成。
2 结果
2.1 取食果实种类
野外共标记637棵119种猴群取食的果树。另有12种植物未在野外发现取食,但在粪便中出现。因此,河静黑叶猴共取食131种果实,分属38科(表1)。
河静黑叶猴取食的果实种类在月间差异显著,多集中在7月至12月间。其中以8月最高,达37种。4月最低,4种。调查期间,在1月至3月未发现河静黑叶猴取食果实。对16833个取食频次的统计结果显示,河静黑叶猴对果实的取食亦集中在7月至12月间(图1),且季节间差异显著,雨季明显高于旱季(z=-2.903,P=0.02<0.05)。
2.2 对果实的选择性
2.2.1 果实性状
131种果实中,以肉质果最多,达106种,其中浆果和核果各53种(各占40.5%),其次是裂果15种,闭果7种,假种皮果实有3种。对果实性状的统计结果显示(表2),河静黑叶猴表现出对黄色果实的明显偏爱,其次是红色和绿色果实(三种颜色的果实共90种,占果实种类的68.7%);取食的果实中能以指甲剥开的为主(共121种,占果实种类的92.4%);偏好取食较大的果实,集中在5g以上(共104种,占果实种类的79.4%)。
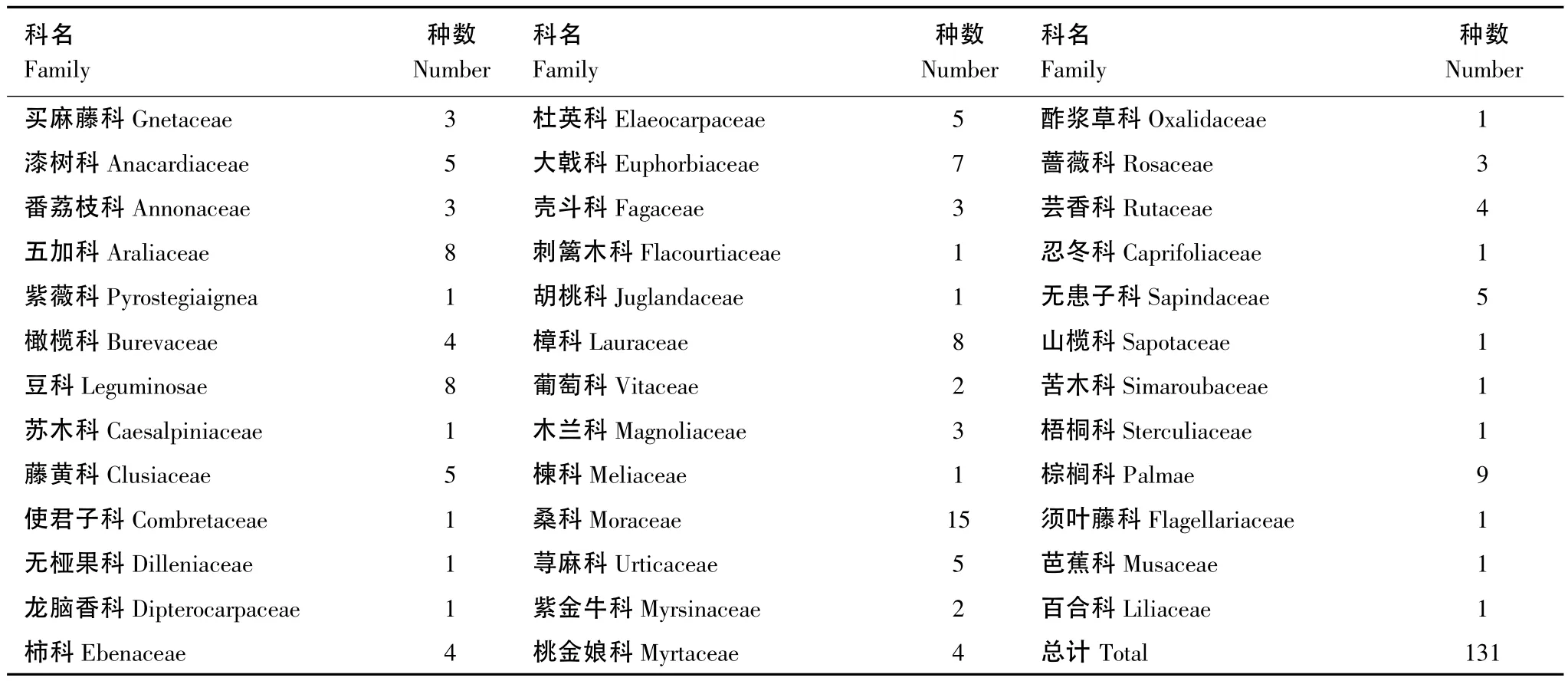
表1 河静黑叶猴取食的果实组成Table 1 Fruit composition consumed by Hatinh langur

图1 河静黑叶猴果实类食物种类及取食频次的月间变化Fig.1 Monthly changes of fruit diet and feeding frequency of Hatinh langurs

表2 河静黑叶猴取食的三种果实性状Table 2 Three characters of fruits consumed by Hatinh langur
河静黑叶猴取食的果实中所含的种子数量差异较大,以1—2颗的为主,有96种,占果实种类的73.3%。果实中种子重量以0.1—1g最多,有57种,占果实种类的43.5%,但≤0.1 g、1—2 g和3—4 g等亦有相当比例的取食(图2)。
2.2.2 果实种类
选择性指数S显示,河静黑叶猴仅对假柿木姜子(Litsea monopetala)(S=0.95)和黄皮 (Clausena lansium)(S=0.84)2种果实具高度选择性;对5种果实,木蝴蝶(Oroxylum indicum)(S= -0.55)、Bauhinia saccocalyx(羊蹄甲属)(S=-0.52)、Garcinia fagraeoides(藤黄属)(S=-0.48)、Clausena laevis(黄皮属)(S=-0.94)和 Licuala fatua(轴榈属)(S=-0.69)表现出低选择性。对其他112种果实呈中度选择性(-0.3<S<0.3)。相关性检验结果发现,河静黑叶猴对果实的取食偏好与果实在生境中的资源可获得性间不相关(r=-0.13,P=0.15>0.05)。
2.3 种子处理与传播
野外共收集粪便2831堆,其中2633(93%)粪便中含有种子,粪便平均包含(4.0±1.5)颗种子。共发现种子16959颗,长度均小于3 cm。经鉴定属85种果实植物。统计结果显示,粪便中绝大多数的种子来自五加科(Araliaceae)、芸香科(Rutaceae)、荨麻科(Urticaceae)、大戟科(Euphorbiaceae)、忍冬科(Caprifoliaceae)、桑科(Moraceae)、龙脑香科(Dipterocarpaceae)、漆树科(Anacardiaceae)和桃金娘科(Myrtaceae)的植物,均为容易处理的浆果或核果(表3)。

图2 河静黑叶猴采食果实的种子数量及重量Fig.2 Seed number and seed weight of fruit consumed by Hatinh langur
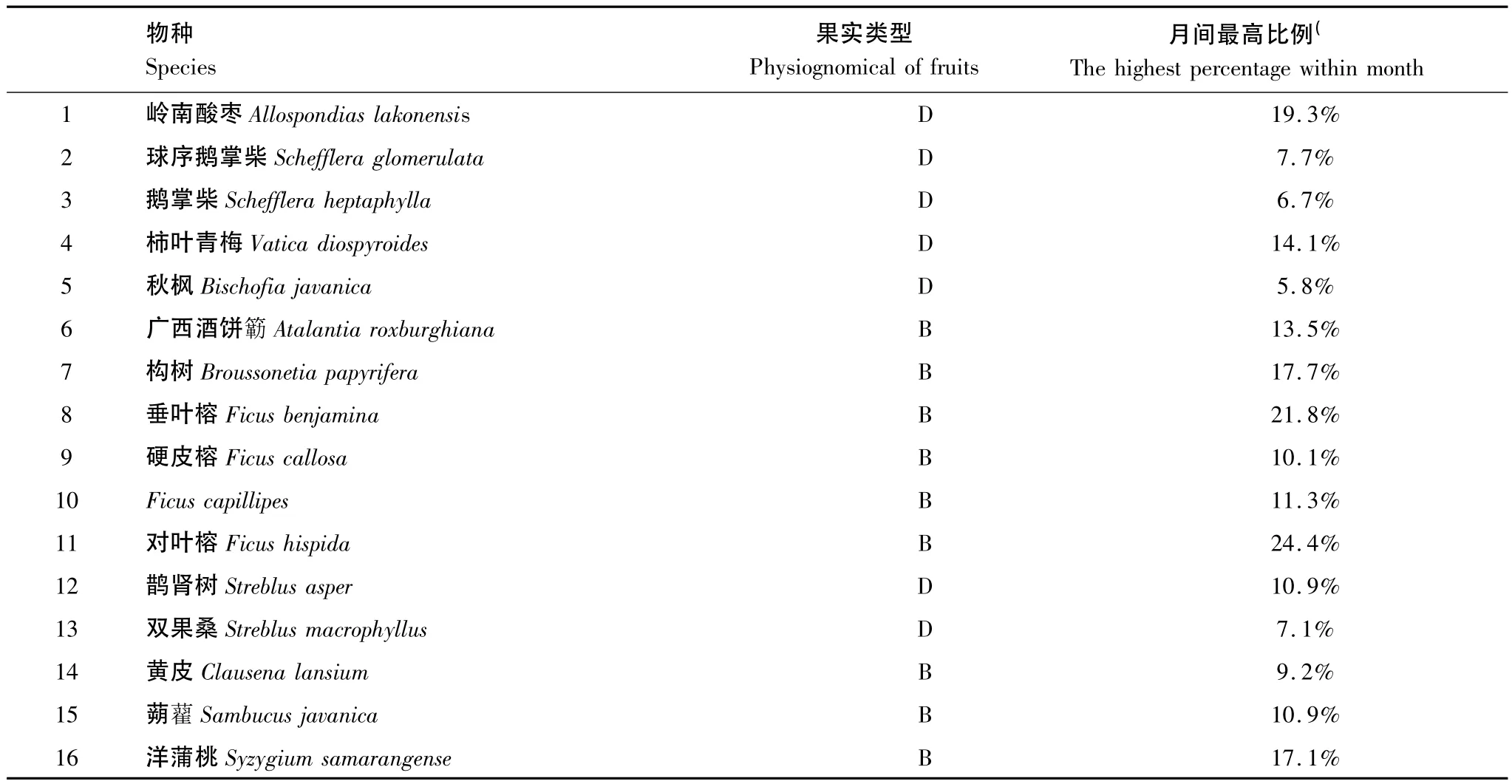
表3 粪便中种子数量最多的16种果实植物Table 3 Top sixteen plants of most numbers of their seeds in feces of monthly records
河静黑叶猴粪便携带种子最近的可在母树下,最远则可达数百米,如九丁榕(Ficus nervosa)(397 m)、垂叶榕(Ficus benjamina)(268 m)、大戟(Bischofia javanica)(247 m)、大参(Macropanax oreophilum)(231 m)、构树(Broussonetia papyrifera)(219 m)和岭南酸枣(Allospondias lakonensis)(107 m)。
河静黑叶猴粪便中种子数量在月份间有一定差异(图3)。2009年和2010年数据显示,8月至12月的夏秋两季是河静黑叶猴粪便中种子数量最多的时期。
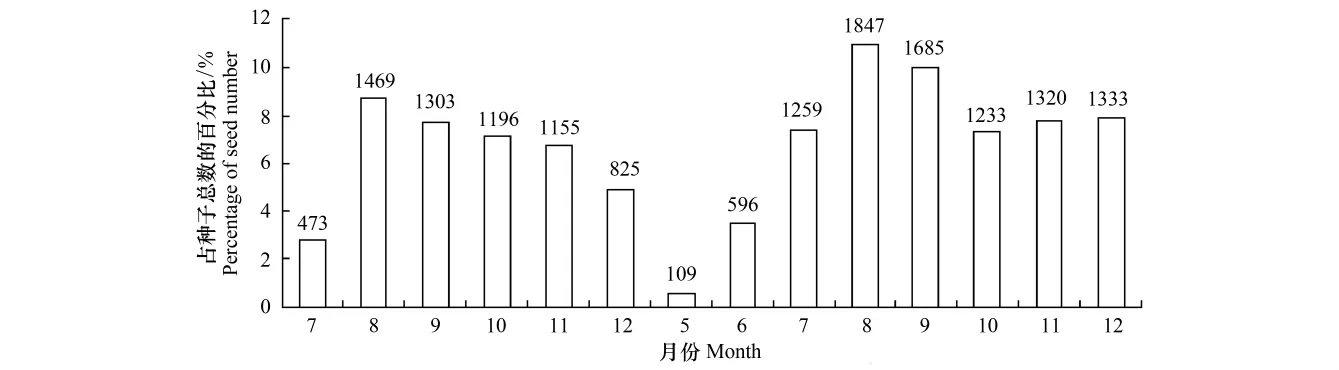
图3 河静黑叶猴传播的种子数量月间比较Fig.3 Comparison among months of seed numbers dispersed by Hatinh langurs
3 讨论
3.1 河静黑叶猴果实类食物及选择
越南的河静黑叶猴全年共采食果实类食物131种,分属38科(表1)。取食的果实种类明显高于其他地区黑叶猴对果实的取食种类,对果实类食物的相对取食高峰期亦长于其他地区(表4)。可能的原因是地区间植被差异及果实类食物多样性不同。有研究证明,生活在连续森林中的灵长类动物比生活在破碎化森林的种类倾向于采食更多的果实[22]。果实的营养高于树叶,但其季节性限制更强,资源量及可获得性较之树叶更低[23],而叶猴对食物的选择受食物的丰富度、质量及分布影响[15]。越南风芽-格邦自然国家公园人为干扰程度小,森林覆盖率93.6%,其中热带季雨性原始森林面积4%,是越南森林覆盖率最高的地区之一。随着纬度的降低,热带雨林中结果植物种类的增加,为黑叶猴对果实的选择多样化提供了条件。

表4 与其他地区黑叶猴果实类食物组成比较Table 4 Comparison of fruit food composition of Francois langur in different sites
河静黑叶猴对果实的取食有季节差异,雨季对果实的取食频次明显高与旱季(Z=-2.903,P=0.02<0.05)。结果反映出河静黑叶猴在果实丰富的雨季能充分利用这类季节性较强的食物(图1),相同的行为亦出现在分布于弄岗地区黑叶猴对果实的取食研究结果[14]。但在对具体的某种果实的取食偏好上却并不受其资源可获得性影响。研究结果显示,河静黑叶猴对果实的取食频次与该果实可获得性的相关性并不显著(r=-0.13,P=0.15>0.05),该结果亦得到选择性指数S的支持,目标猴群对112种果实呈中度选择,反映出河静黑河后对果实的取食偏好与该果实在环境中的可获得性关系不大。虽然本项调查仅以果实类食物为研究对象,但分析结果与其他叶猴食性研究相似。如长尾叶猴(Presbytis entellus)取食的28种食物中,对其中25种食物的选择没有受到食物资源量的影响[25];黑乌叶猴(T.johnii)共取食115种植物,但它们并不偏好更多地取食资源量大的食物,包括果实类食物[26];广西白头叶猴(T.leucocephalus)对食物的选择与食物资源的可获得性亦不相关[23,]。由此可见,河静黑叶猴对果实食物的选择偏好可能更易受到果实性状以及自身的营养需求影响。这方面还需进一步的验证。
在果实性状上,河静黑叶猴对处理难度较小的浆果和核果有明显偏好(表3),它们更多地选择黄色、红色和绿色,重量多为5g以上较大的果实,更多地选择果实内仅有1—2颗种子的果实(表2,图2)。河静黑叶猴对果实性状的选择偏好反映了灵长类对果实选择的一般特点,如主要取食果皮容易处理的肉质果,多为黄色、红色、褐色或绿色,以大中型果实为主,多数在5 g至50 g[6,7]。大多数灵长类喜好大型肉质果[6],果实越大,种子越少,可以消化的果肉就越多;越容易处理,则单位时间内所获取的净收益越大[27]。对果实颜色的选择可能与河静黑叶猴发现果实的难易程度有关。一般来说,黄色和红色的果实与树叶背景反差很大,能更容易地吸引到灵长类动物的注意[28]。野外调查发现,河静黑叶猴取食的果实中,以成熟果实为多,有96种,占72.3%,不成熟果实35种,占27.7%。成熟的鲜艳果实意味着含有更多的糖分和能量[21,29]。这与前人对疣猴相关方面的研究结果不同。有研究发现过多地摄入糖分会改变疣猴胃内的pH值,进而影响胃内微生物的发酵活动以消化树叶,如弄岗黑叶猴大部分采食未成熟的果实[15]。除了长期适应于当地食物环境外,食物所含化学成分与动物个体对营养成分的消化率差异亦是可能的原因。如在对分布于印度和尼泊尔的两个长尾叶猴(Semnopithecus entellus)种群的食物营养利用比较研究发现,两个种群间对营养成分的消化率差异显著。相对于尼泊尔种群,印度种群的个体对蛋白质和糖分的消化率更低。作者认为种群或个体对营养成分的消化率差异在满足自身营养需求和食物选择中,比食物本身具有更重要的作用[30]。实际上大多数灵长类动物对果实颜色的选择范围均较广[31-32]。它们对果实特征的选择除了与生境中所能提供的果实种类及其资源量[8]、行为模式[9,33]不同有关外,还与动物的生理特征[11,22]和营养需求[34]有关。
3.2 河静黑叶猴对喀斯特森林中种子扩散的意义
河静黑叶猴对种子的处理方式主要有三种,(1)大且硬的种子,多在取食过程中吐出。或随啃食一半的果实丢弃,致使种子掉落地点围绕在母树周围,距离较近,多集中在6—20 m范围,如云南山竹子(Garcinia merguensis)、岭南山竹子(Garcinia oblongifolia)、杧果(Mangifera indica)和山杜英(Elaeocarpus limitaneus)等植物;(2)部分小且种皮较软的种子常被嚼烂,如银柴(Aporosa dioica)、辛果漆(Drimycarpus racemosus)和五倍子(Rhus chinensis)等小果实植物;(3)许多小种子的浆果和核果,如桑科(Moraceae)和大戟科(Euphorbiaceae)植物,整吞后以消化道的方式远距离携带,然后随粪便排出。野外观察发现,河静黑叶猴对很多果实同时存在三种种子处理方式,如桑科植物的很多浆果,取食过程中,除了吞食种子外,还有相当数量的种子被吐出或随啃食一半的果实散落在母树周围,在粪便里还发现了这些种子被嚼烂后的种皮碎片。
河静黑叶猴对喀斯特森林中的果实植物具有潜在的种子传播作用。研究结果显示,有多达85种果实的种子能被河静黑叶猴吞食,再通过动物的运动将种子传播至较远的适宜萌发生境,尤以种子直径小于3cm的浆果和核果最多,如垂叶榕、柿叶青梅(Vatica diospyroides)、构树、岭南酸枣、洋蒲桃(Syzygium samarangense)和对叶榕(Ficus hispida)等(表3)。最远传播距离可达397 m(九丁榕)。对另外的46种果实,河静黑叶猴则以吐出或丢弃等方式短距离传播。在传播功能上,一些果实较大或果皮较硬的植物,由于鸟类无法吞食其果实,如水浦桃(Syzygium jambos)、红毛丹 (Nephelium lappaceum)和木奶果(Baccaurea ramiflora)等,河静黑叶猴可能是其潜在的种子传播者。
灵长类的种子传播对热带森林维持和演化具重要影响[35],它们数量众多,对果实的取食量大,传播距离远,很多植物都需要依靠灵长类动物来传播种子[6,9,36]。喀斯特森林是一类特殊生境,生态环境脆弱,具有植物多样性高但资源量低的特点[13-14],植物的种子扩散与更新对维持森林生态系统极其重要。河静黑叶猴能够通过肠道远距离携带85种植物的种子,且一年中超过6个月取食果实,尤其在雨季的8月至12月间对果实的取食强度达到高峰,这期间猴群粪便中的种子数量均在1000以上(图3),显示了较强的种子潜在传播功能。本项研究初步了解了河静黑叶猴对果实类食物组成、对果实特征的选择以及潜在的种子传播作用,更深入的种子传播机制以及河静黑叶猴与果实植物间的互惠共生关系如何影响喀斯特森林的植被组成与演化还需进一步的研究。
[1] Oates J F.The natural history of African colobines//Davies A G,Oates J F eds.Colobine Monkeys:Their Ecology,Behaviour and Evolution.UK:Cambridge University Press,1994:75-128.
[2] Yeager C P,Kool K.The Behavioral Ecology of Asian Colobines//Whitehead P F,Jolly C J,eds.Old World Monkeys.UK:Cambridge University Press,2000:497-521.
[3] Bennett E L.The Banded Langur:Ecology of A Colobine in A West Malaysian Rain Forest[D].England:University of Cambridge,1983.
[4] Gurmaya K J.Ecology and behavior of Presbytis thomasi in Northern Sumatra.Primates,1986,27(2):151-172.
[5] Li Z Y,Rogers M E.Food items consumed by white-headed Langurs in Fusui,China.International Journal of Primatology,2006,27(6):1551-1567.
[6] Gautier-Hion A,Duplantier J M,Quris R,Quris R,Feer F,Sourd C,Decoux J P,Dubost G,Emmons L,Erard C,Hecketsweiler P,Moungazi A,Roussihon C,Thiollay J M.Fruit characters as a basis of fruit choice and seed dispersal in a tropical forest vertebrate community.Oecologia,1985,65(3):324-337.
[7] Janson C H.Adaptation of fruit morphology to dispersal agents in a neo-tropical forest.Science,1983,219(4581):187-189.
[8] Lambert J E,Chapman C A,Wrangham R W,Conklin-Brittain N L.Hardness of cercopithecine foods:implications for the critical function of enamel thickness in exploiting fallback foods.American Journal of Physical Anthropolog,2004,125(4):363-368.
[9] McConkey K R.Primary seed shadow generated by gibbons in the rain forests of Barito Ulu,central Borneo.American Journal of Primatology,2000,52(1):13-29.
[10] Riba-Hernández P,Stoner K E,Osorio D.Effect of polymorphic colour vision for fruit detection in the spider monkey Ateles geoffroyi,and its implications for the maintenance of polymorphic colour vision in platyrrhine monkeys.The Journal of Experimental Biology,2004,207(14):2465-2470.
[11] Birkinshaw C.Fruit characteristics of species dispersed by the black Lemur(Eulemur macaco)in the Lokobe Forest,Madagascar.Biotropica,2001,33(3):478-486.
[12] Yao C,Jiang Z C,Yuan D.Vegetation Karst effects on the karst area of southwest China.Acta Geoscientia Sinica,2001,22(2):159-164.
[13] Zhou Q H,Cai X W,Huang B M,Li Y B,Luo Y P.Feeding activity of Francois langurs in Karst habitat.Acta Theriologica Sinica,2007,27(3):243-248.
[14] Huang Z H,Huang B M,Zhou Q H,Wei H,Meng Y J.Diet and the seasonal changes of the Fran ois'langur(Trachypithecus francoisi).Acta Ecologica Sinica,2010,30(20):5501-5508.
[15] Zhou Q H,Wei F W,Li M,Huang C M,Luo B.Diet and food choice of Trachypithecus francoisi in the Nonggang nature reserve,China.International Journal of Primatology,2006,27(5):1441-1459.
[16] Hu G.Dietary breadth and resource use of Franc-ois'langur in a seasonal and disturbed habitat.American Journal of Primatology,2011,73(11):1176-1187.
[17] Haus T,Vogt M,Forster B,Thanh Vu N,Ziegler M.Distribution and Population Densities of Diurnal Primates in the Karst Forests of Phong Nha-Ke Bang National Park,Quang Binh Province,Central Vietnam.International Journal of Primatology,2009,30(2):301-312
[18] Nguyen M H.Some Observations on the Hatinh langur,Trachypithecus laotum hatinhensis(Dao,1970),in North Central Vietnam.Primate Conservation,2006,21:149-154
[19] Altmann J.Observational study of behavior:Sampling methods.Behaviour,1974,49(3):227-267.
[20] Julliot C,Sabatier D.Diet of the red Howler monkey(Alouatta seniculus)in French Guiana.International Journal of Primatology,1993,14(4):527-550.
[21] Sourd C,Gautier-Hion A.Fruit selection by a forest guenon.Journal of Animal Ecology,1986,55(1):235-244.
[22] Tutin C E G,Ham R M,White L J T,Harrison M J S.The primate community of the Lop Reserve,Gabon:diets,responses to fruit scarcity,and effects on biomass.American Journal of Primatology,1997,42:1-24.
[23] Li Z Y,Wei Y,Rogers E.Food choice of white-headed Langurs in Fusui,China.International Journal of Primatology,2003,24(6):1189-1205.
[24] Cai X W.The research on feeding biology and nutrition analysis of Presbytis francoisi[D].Guilin:Guangxi Normal University,2004.
[25] Hladik C M A.A comparative study of the feeding strategies of two sympatric species of leaf monkeys:Presbytis senex and Presbytis entellus//Clutton-Brock T H,ed.Primate Ecology:Studies of Feeding and Ranging Behaviour in Lemurs,Monkeys and Apes.London:Academic Press,1977:323-353.
[26] Oates J F,Waterman P G,Choo G M.Food selection by the south Indian leaf-monkey,Presbytis johnii,in relation to leaf chemistry.Oecologia,1980,45(1):45-56.
[27] Chapman C A.Primate seed dispersal:the fate of dispersed seeds.Biotropica,1989,21(2):148-154.
[28] Párraga C A,Troscianko T,Tolhurst D J.Spatiochromatic properties of natural images and human vision.Current Biology,2002,12(6):483-487.
[29] Wheelwright N T,Janson C H.Colors of fruit displays of bird-dispersed plants in two tropical forests.American Naturalist,1985,126(6):777-799.
[30] Schulke O,Chalise M,Koenig A.The importance of ingestion rates for estimating food quality and energy intake.American Journal of Primatology,2006,68(10):951-965.
[31] Dominy N J,Lucas P W.The visual ecology of catarrhines and the role of color,calories,and climate.American Journal of Primatology,2004,62(3):189-207.
[32] Stoner K E,Riba-hernandez P,Lucas P W.Comparative use of colorvision for frugivory by sympatric species of Platyrrhines.American Journal of Primatology,2005,67(4):399-409.
[33] Stevenson P R.Seed dispersal by wooly monkeys(Lagothrix lagothricha)at Tinigua National park,colombia:dispersal distance,germination rates and dispersal quantity.American Journal of Primatology,2000,50(4):275-289.
[34] Wasserma M D,Chapman C A.Determinants of colobine monkey abundance:the importance of food energy,protein and fibre content.Journal of Animal Ecology,2003,72(4):650-659.
[35] Chapman C A.Primate seed dispersal:coevolution and conservation implications.Evolutionary Anthropology,1995,4(3):74-82.
[36] Lambert J E.Seed handling in chimpanzees(Pan troglodytes)and redtail monkeys(Cercopithecus ascanius):implications for understanding hominoid and cercopithecine fruit-processing strategies and seed dispersal.American Journal of Physical Anthropology,1999,109(3):365-386.
参考文献:
[12] 黄乘明.中国白头叶猴.桂林:广西师范大学出版社,2002.
[13] 周岐海,蔡湘文,黄乘明,李友邦,罗亚平.黑叶猴在喀斯特石山生境的觅食活动.兽类学报,2007,27(3):243-248.
[14] 黄中豪,黄乘明,周岐海,韦华,蒙渊君.黑叶猴食物组成及其季节性变化.生态学报,2010,30(20):5501-5508.
[24] 蔡湘文.黑叶猴的觅食生物学和营养分析[D].桂林:广西师范大学,2004.
猜你喜欢
发明与创新(2021年45期)2021-02-16
第二课堂(课外活动版)(2021年10期)2021-02-13
环球时报(2020-07-22)2020-07-22
小学阅读指南·低年级版(2019年3期)2019-05-13
收藏界(2018年1期)2018-10-10
中国周刊(2018年6期)2018-06-15
创新作文(小学版)(2018年31期)2018-05-16
Zoological Research(2017年4期)2017-08-24
摄影之友(影像视觉)(2017年1期)2017-07-18
小学阅读指南·低年级版(2016年6期)2016-05-14
