
蒙古栎地理分布的主导气候因子及其阈值
2013-12-19 07:30殷晓洁周广胜2隋兴华何奇瑾李荣平
生态学报 2013年1期
殷晓洁,周广胜2,,* ,隋兴华,何奇瑾,李荣平
(1.中国科学院植物研究所植被与环境变化国家重点实验室,北京 100093;2.中国气象科学研究院,北京 100081;3.南京信息工程大学大气物理学院,南京 210044;4.中国气象局沈阳大气环境研究所,沈阳 110016)
人类活动引起的大气温室气体增加导致的气候变暖,将影响到降水、辐射、潜在蒸散等其他气候变量发生变化,进而影响生态系统,特别是物种分布及植被带的迁移[1]。
蒙古栎(Quercus mongolica),又称柞木、柞树,是东亚—东西伯利亚分布种,在我国主要分布于东北地区和华北地区,是我国温带地区落叶阔叶林及针阔混交林的主要树种[2-5]。研究表明,气候是区域尺度上决定物种分布的主要因子[1],未来气候变化将使蒙古栎地理分布范围扩大,成为我国大兴安岭和小兴安岭最主要树种[6]。但是,目前关于影响蒙古栎地理分布的主导气候因子及其阈值的研究还较少,制约着蒙古栎对气候变化响应的理解,也影响着蒙古栎林的科学经营管理。
通常,采用物种分布模型进行物种的地理分布及其对气候变化响应的模拟研究。物种分布模型(species distribution models)主要是基于物种的已知分布及其环境变量,探索物种的生态位和潜在分布。目前,物种分布模型已经广泛用于物种潜在分布区的预测,包括生态位模型(BIOCLIM、BLOMAPPER、DIVA、DOMAIN)、动态模拟模型(CLIMEX)、广义相加模型GAM、广义线性模型GLM、基于检验假设的分布预测模型GARP(the genetic algorithm for rule-set prediction)以及最大熵模型(Maxent)等[7-11],不同模型的侧重点不同[12-20]。其中,最大熵模型被许多研究证明是对物种分布具较好预测能力的模型[7-8,21-33]。
本研究试图以蒙古栎为研究对象,基于气候相似性原理,利用Maxent模型筛选影响蒙古栎地理分布的主导气候因子,给出其分布范围及各主导影响因子的阈值,并进行其气候适应性划分,以为蒙古栎林的经营管理及应对气候变化提供依据。
1 资料与方法
1.1 蒙古栎地理分布资料
蒙古栎地理分布资料通过3个途径获取:(1)《中华人民共和国植被分布图(1∶1000000)》的蒙古栎分布资料;(2)中国科学院植物研究所标本馆提供的标本数字资料;(3)各地植物志的蒙古栎分布资料,包括《北京植物志》、《河北植物志》、《山东植物志》、《河南植物志》、《崂山植物志》、《黄土高原植物志》、《黑龙江植物志》、《辽宁植物志》、《内蒙古植物志》等。提取《中华人民共和国植被分布图(1∶1000000)》中蒙古栎各分布区的几何中心点坐标,结合标本馆标本数字资料和植物志的蒙古栎各分布区几何中心点坐标,共同构成我国蒙古栎地理分布数据集。利用3个途径获取蒙古栎地理分布资料时,如有重叠,按《中华人民共和国植被分布图(1∶1000000)》、标本馆标本数字资料和各地植物志的顺序选取。
1.2 气象资料
气象数据来自中国气象局国家气象信息中心的国家基本气象观测台站(756个),包括1971—2000年的日均温度和日降水量等。采用空间卷积原理和截断高斯滤波算子方法,结合各气象站地理信息差值得到中国10 km×10 km空间分辨率的日均气温和日降水量数据[34-35]。
1.3 影响蒙古栎地理分布的潜在气候因子选取
Woodward指出,影响植物地理分布的主要因子有三类:(1)植物的耐寒性;(2)完成生活史所需的生长季长度和热量供应;(3)用于植物冠层形成和维持的水分供应[36]。据此,本研究选取一月均温(Tc)作为最冷月温度,反映植物的耐寒性;用7月均温(Tm)作为最暖月温度,与大于5℃积温(GDD5)共同反映热量需求;采用年均降水量(P)和湿润指数(MI,年降水量和年潜在蒸散量的比值=P/PET)表示水分需求。同时,选取气温年较差(DTY)反映气温变幅。年潜在蒸散(PET)采用Thornthwaite方法计算[37-39]。
1.4 研究方法
Maxent模型以最大熵理论为基础,根据不完全信息,从符合条件的分布中选择熵最大的分布作为最优分布,建立预测模型,进而预测物种的地理分布[7,21]。模型基于贝叶斯定理,利用Gibbs分布族将特征集进行加权并作为参数,进行一系列运算,得到物种分布的最大熵联合分布估计而建立[7]。Maxent模型被证明对物种分布有非常好的预测能力并具有很多优点[7,8,21-24],如简单而清晰的数学基础,易于从生态学上进行解释;连续型和分类型的环境变量都可以使用;只需要模拟物种的当前存在数据等。
本研究选取Maxent模型进行蒙古栎气候适应性研究。首先,确定特征空间,即物种已知分布区域;其次,寻找限制物种分布的约束条件(潜在气候因子),构筑约束集合;第三,利用Maxent模型构建蒙古栎地理分布与气候关系模型[7],并检验模型的适用性;第四,基于各潜在气候因子对蒙古栎地理分布的贡献,筛选影响我国蒙古栎地理分布的主导气候因子,重新构建基于Maxent模型的蒙古栎地理分布与气候关系模型;第五,基于我国蒙古栎地理分布给出影响我国蒙古栎地理分布主导气候因子的阈值。
其中,采取常用的ROC曲线(受试者工作特征曲线)下的面积即AUC值[40-41]作为模型预测准确性的衡量指标(表1),以检验模型的适用性。通过模型Jackknife模块分析各潜在气候因子对蒙古栎分布模拟的得分情况。基于ArcGIS地理信息系统,得到Maxent模型模拟的蒙古栎地理分布。

表1 AUC值及其与模型准确性的关系[41]Table 1 Relationship between AUC and the accuracy of the model
2 结果分析
2.1 模型适用性分析
为验证基于最大熵模型构建的蒙古栎地理分布与气候关系模型对我国蒙古栎地理分布研究的适用性,首先需要基于训练子集(对整个数据随机取样取得总数据集的75%作为训练子集)来训练模型,获取模型的相关参数,构建针对我国蒙古栎地理分布的最大熵模型;然后,将没有参与模型构建的所有数据用作评估子集(即余下数据的25%),用来验证模型。模型运行需要两组数据,一是目标物种的地理分布数据,即构建的我国蒙古栎地理分布数据集;二是全国范围的气候变量,即基于已有研究成果从全国层次及年尺度筛选出的6个潜在气候因子。
采用AUC值作为模型预测准确性的衡量指标。结果表明,基于潜在气候因子与最大熵模型构建的我国蒙古栎地理分布与气候关系模型的AUC值达0.932,表明所构建模型的预测准确性达到“极准确”的水平,可以用于我国蒙古栎地理分布与气候的关系研究。
2.2 影响我国蒙古栎地理分布的主导气候因子分析
选取的潜在气候因子主要来源于已有的研究成果,而没有从全国层次上定量评价这些因子对蒙古栎地理分布的影响程度,这将影响其最大熵模型构建变量的准确选择,进而影响到我国蒙古栎地理分布模拟的准确性。为此,需要定量评价这些潜在气候因子对我国蒙古栎地理分布的影响程度,筛选主导气候因子。
图1是基于最大熵模型的Jackknife模块给出的各潜在气候因子对我国蒙古栎地理分布影响的得分情况,以此反映各因子的贡献。其中,图中横轴表示因子得分值,纵轴表示各因子,深蓝色棒、浅蓝色棒和底部的红色棒分别表示该因子得分、除该因子外其它指标得分之和及所有指标的得分和。各潜在气候因子对我国蒙古栎地理分布影响的贡献排序为:年均降水量(P)>气温年较差(DTY)>大于5℃有效积温(GDD5)>最暖月温度(Tw)>湿润指数(MI)>最冷月温度(Tc)。
结果表明,最冷月温度和湿润指数对我国蒙古栎地理分布的贡献较低,与蒙古栎具有较强的耐寒性和耐旱性相一致[5,42],可以不作为主导气候因子。根据各潜在气候因子对我国蒙古栎地理分布影响的贡献情况结合蒙古栎生理生态特性[5,42-43],确定影响我国蒙古栎地理分布的主导气候因子为:年均降水量、气温年较差、大于5℃积温和最暖月温度,这4个因子的百分贡献率累积值达92%。

图1 各气候因子Jackknife检验得分Fig.1 Jackknife test gain of each climate factor
2.3 影响我国蒙古栎地理分布的主导气候因子阈值分析
基于最大熵模型以及选定的影响我国蒙古栎地理分布的4个主导气候因子,可以给出蒙古栎在预测地区的存在概率P,取值范围为0—1。由统计学原理可知,当植物在某一地区的存在概率P<0.05时,其出现的概率很小,即小概率事件,在此定义该植物在该地区不能存在。为分析影响我国蒙古栎地理分布的主导气候因子阈值,首先需要弄清所建模型对我国蒙古栎地理分布模拟的准确性。为此,采用最大熵模型确定的影响我国蒙古栎地理分布的4个主导气候因子:年均降水量、气温年较差、大于5℃积温和最暖月温度,以模型给出的蒙古栎存在概率0.05为界,给出蒙古栎在我国的地理分布(图2),并与蒙古栎实际地理分布进行比较。
结果表明,蒙古栎在黑龙江、吉林、辽宁、内蒙古东部、北京、天津、河北中北部、山东中部、山西大部分地区和陕西东北部均可分布。该区域覆盖了蒙古栎实际地理分布的1013个分布点中的1002个,覆盖率大于98%,表明模拟的我国蒙古栎地理分布与实际分布非常符合。

图2 Maxent模型模拟的蒙古栎分布Fig.2 Distribution of Q.mongolica simulated by Maxent model
采用最大熵模型确定的影响我国蒙古栎地理分布的4个主导气候因子:年均降水量、气温年较差、大于5℃积温和最暖月温度,结合我国蒙古栎地理分布,从蒙古栎地理分布概率与主导气候因子的关系可以给出各主导气候因子的阈值(模型的训练重复数设为10次)。由图3可见,较粗的蓝线表示多次重复得到的阈值,红线表示多次重复阈值的均值,从而得到影响我国蒙古栎地理分布主导气候因子的阈值为:年均降水量为330—910 mm、气温年较差大于29℃、大于5℃有效积温为1200—3500℃·d、最暖月均温为17—26℃。气温年较差要求大于29℃表明,蒙古栎林适合生长在高纬度或高海拔地区;最暖月温度要求17—26℃表明蒙古栎适合生长在夏季温度偏低地区;年均降水量和大于5℃积温限定了东北地区、华北地区、西北部分地区以及山东省和湖北省为符合蒙古栎林分布的水分和热量条件区。
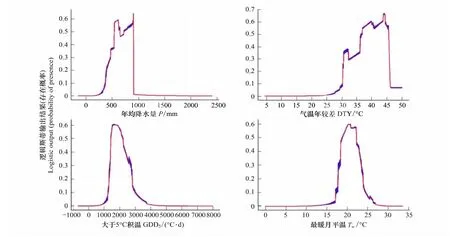
图3 影响我国蒙古栎地理分布的主导气候因子与蒙古栎分布概率的关系Fig.3 Relationship of each dominant climatic factor and the distribution probability of Q.mongolica
3 结论
基于我国蒙古栎地理分布信息,结合1971—2000年我国10 km×10 km空间分辨率的气候资料及影响我国蒙古栎地理分布的潜在气候因子,从气候的相似性出发,验证了利用最大熵模型(Maxent)构建的蒙古栎地理分布与气候关系模型的适用性。结果表明,基于潜在气候因子与最大熵模型构建的我国蒙古栎地理分布与气候关系模型的AUC值达0.932,表明所构建模型的预测准确性达到“极准确”的水平,可以用于我国蒙古栎地理分布与气候关系研究。
根据各潜在气候因子对蒙古栎潜在地理分布的贡献,筛选出了影响蒙古栎分布的主导气候因子,即年均降水量、气温年较差、最暖月温度和大于5℃积温。利用我国蒙古栎地理分布概率与主导气候因子的关系给出了各主导气候因子的阈值,即:年均降水量为330—910 mm、气温年较差大于29℃、大于5℃有效积温为1200—3500℃·d,最暖月均温为17—26℃。该研究有助于增进蒙古栎与气候变化关系的理解,对于科学地经营和管理蒙古栎及制定应对气候变化对策具有重要意义。
[1] Zhou G S,Wang Y H.Global Change Ecology.Beijing:Meteorological Press,2003:119-122.
[2] Wu Z Y.Vegetation of China.Beijing:Science Press,1980.
[3] Wang L M,Ren X W,Liu Y J.Geographic distribution of deciduous oaks in China.Journal of Beijing Forestry University,1985,(2):57-69.
[4] Zheng H N,Jia S Q,Hu H Q.Forest fire and forest rehabilitation in the DA XINGAN mountains.Journal of Northeast Forestry University,1986,14(4):1-7.
[5] Yu S L,Ma K P,Chen L Z.Preliminary discussion on the origin of Quercus mongolica forest in north China.Guihaia,2000,5(2):131-137.
[6] Yan X D,Zhao S D,Yu Z L.Modeling growth and succession of Northeastern China forests and its applications in global change studies.Journal of Plant Ecology,2000,24(1):1-8.
[7] Phillips S J,Dudik M.Modeling of species distributions with Maxent:new extensions and a comprehensive evaluation.Ecography,2008,31:161-175.
[8] Elith J,Graham C H,Anderson R P,Dudík M,Ferrier S,Guisan A,Hijmans R J,Huettmann F,Leathwick J R,Lehmann A,Li J,Lohmann L G,Loiselle B A,Manion G,Moritz C,Nakamura M,Nakazawa Y,Overton J M M,Peterson A T,Phillips S J,Richardson K,Scachetti-Pereira R,Schapire R E,Soberón J,Williams S,Wisz M S,Zimmermann N E.Novel methods improve prediction of species'distributions from occurrence data.Ecography,2006,29(2):129-151.
[9] Kriticos D,Randall R.A comparison of systems to analyse potential weed distributions//Grove R H,Panetta F D,Virtue J G,eds.Weed Risk Assessment.Australia:CSRIO Publishing,2001:61-79.
[10] Guisan A,Thuiller W.Predicting species distribution:offering more than simple habitat models.Ecology Letters,2005,8(9):993-1009.
[11] Sun W T,Liu Y T.Research progress of risk analysis of biological invasion.Chinese Agricultural Science Bulletin,2010,26(7):233-236.
[12] Carpenter G,Gillson A N,Winter J.DOMAIN:a flexible modeling procedure for mapping potential distributions of plants and animals.Biodiversity and Conservation,1993,2(6):667-680.
[13] Prentice I C,Cramer W,Harrison S P,Leemans R,Monserud R A,Solomon A M.A global biome model based on plant physiology and dominance,soil properties and climate.Journal of Biogeography,1992,19(2):117-134.
[14] Guisan A,Zimmermann N E.Predictive habitat distribution models in ecology.Ecological Modelling,2000,135(2/3):147-186.
[15] Vayssières M P,Plant R E,Allen-Diaz B H.Classification trees:an alternative non-parametric approach for predicting species distributions.Journal of Vegetation Science,2000,11(5):679-694.
[16] Cao M C,Zhou G S,Weng E S.Application and comparison of generalized models and classification and regression tree in simulating tree species distribution.Acta Ecologica Sinica,2005,25(8):2031-2040.
[17] Li F,Zhou G S,Cao M C.Responses of Larix gmelinii geographical distribution to future climate change:a simulation study.Chinese Journal of Applied Ecology,2006,17(12):2255-2260.
[18] Wang J,Ni J.Review of modeling the distribution of plant species.Journal of Plant Ecology,2006,30(6):1040-1053.
[19] Iverson L R,Prasad A M.Potential redistribution of tree species habitat under five climate change scenarios in the eastern US.Forest Ecology and Management,2002,155(1/3):205-222.
[20] Thuiller W,Araújo M B,Lavorel S.Generalized models versus classification tree analysis:predicting spatial distributions of plant species at different scales.Journal of Vegetation Science,2003,14(5):669-680.
[21] Phillips S J,Anderson R P,Schapire R E.Maximum entropy modeling of species geographic distributions.Ecological Modelling,2006,190(3/4):231-259.
[22] Ma S M,Zhang M L,Zhang H X,Meng H H,Chen X.Predicting potential geographical distributions and patterns of the relic plant Gymnocarpos przewalskii using Maximum Entropy and Genetic Algorithm for Rule-set Prediction.Chinese Journal of Plant Ecology,2010,34(11):1327-1335.
[23] Lei J C,Xu H G.Maxent-based prediction of potential distribution of Solidago canadensis in China.Journal of Ecology and Rural Environment,2010,26(2):137-141.
[24] Wang Y S,Xie B Y,Wan F H,Xiao Q M,Dai L Y.Potential geographic distribution of radopholus similis in China.Scientia Agricultura Sinica,2007,40(11):2502-2506.
[25] Giovanelli J G R,Haddad C F B,Alexandrino J.Predicting the potential distribution of the alien invasive American bullfrog(Lithobates catesbeianus)in Brazil.Biological Invasions,2008,10(5):585-590.
[26] Moffett A,Shackelford N,Sarkar S.Malaria in Africa:vector species'niche models and relative risk maps.PLoS ONE,2007,2(9):e824,doi:10.1371/journal.pone.0000824.
[27] Saatchi S,Buermann W,ter Steege H,Morid S,Smith T B.Modeling distribution of Amazonian tree species and diversity using remote sensing measurements.Remote Sensing of Environment,2008,112(5):2000-2017.
[28] Cao X F,Qian G L,Hu B S,Liu F Q.Prodiction of potential suitable distribution area of Flaveria bidentis in China based on niche models.Chinese Journal of Applied Ecology,2010,21(12):3063-3069.
[29] Yang B,Xue Y G,Tang X F,Wang Y,Zhang D M.Predicting potential geographic distribution of Eupatorium odoratum in China.Plant Protection,2009,35(4):70-73.
[30] Wu W H,Li M Y.Methods for predicting potential habitat of bursaphelenchus xylophilus based on ecological niche model.Forest Inventory and Planning,2009,34(5):33-38.
[31] Huntley B,Berry P M,Cramer W,McDonald A P.Modeling present and potential future ranges of some European higher plants using climate response surfaces.Journal of Biogeography,1995,22(6):967-1001.
[32] Berry P M,Dawson T E,Harrison P A,Pearson R G.Modelling potential impacts of climate change on the bioclimatic envelope of species in Britain and Ireland.Global Ecology and Biogeography,2002,11(6):453-462.
[33] Franklin J.Predicting the distribution of shrub species in southern California from climate and terrain-derived variables.Journal of Vegetation Science,1998,9(5):733-748.
[34] Thornton P E,Running S W,White M A.Generating surfaces of daily meteorological variables over large regions of complex terrain.Journal of Hydrology,1997,190(3/4):214-251.
[35] Liu Y,Chen P Q,Zhang W,Hu F.A spatial interpolation method for surface air temperature and its error analysis.Chinese Journal of Atmospheric Sciences,2006,30(1):146-152.
[36] Woodward FI.Climate and Plant Distribution.Cambridge:Cambridge University Press,1987.
[37] Weng E S,Zhou G S.Defining plant functional types in China for global change studies.Acta Phytoecologica Sinica,2005,29(1):81-97.
[38] Zhang X S.The potential evapotranspiration(PE)index for vegetation and vegetation-climatic classification(2)an introduction of main methods and PEP program.Journal of Plant Ecology,1989,13(3):197-207.
[39] Ni J.Relationship between Thornthwaite indexes and distribution of dominants and companions of subtropical evergreen broadleaved forest in China.Plant Science Journal,1997,15(2):151-161.
[40] Chen P F,Wiley E O,Mcnyset KM.Ecological niche modeling as a predictive tool:silver and bighead carps in North America.Biological Invasions,2006,9(1):43-51.
[41] Shao H,Tian J Q,Guo K,Sun J X.Effects of sample size and species traits on performance of BIOCLIM in predicting geographical distribution of tree species-a case study with 12 deciduous Quercus species indigenous to China.Chinese Journal of Plant Ecology,2009,33(5):870-877.
[42] Xu ZQ,Wang YH.Research advances in Quercus mongolica.Hebei Journal of Forestry and Orchard Research,2002,17(4):365-370.
[43] Guo JP,Gao SH,Liu L,Zhou G S.Impacts of climatic change on Quercus mongolica.Resources Science,2005,27(5):168-172.
参考文献:
[1] 周广胜,王玉辉.全球生态学.北京:气象出版社,2003:119-122.
[2] 吴征镒.中国植被.北京:科学出版社,1980.
[3] 王良民,任宪威,刘一樵.我国落叶栎的地理分布.北京林学院学报,1985,(2):57-69.
[4] 郑焕能,贾松青,胡海清.大兴安岭林区的林火与森林恢复.东北林业大学学报,1986,14(4):1-7.
[5] 于顺利,马克平,陈灵芝.中国北方蒙古栎林起源和发展的初步探讨.广西植物,2000,5(2):131-137.
[6] 延晓冬,赵士洞,于振良.中国东北森林生长演替模拟模型及其在全球变化研究中的应用.植物生态学报,2000,24(1):1-8.
[11] 孙文涛,刘雅婷.生物入侵风险分析的研究进展.中国农学通报,2010,26(7):233-236.
[16] 曹铭昌,周广胜,翁恩生.广义模型及分类回归树在物种分布模拟中的应用与比较.生态学报,2005,25(8):2031-2040.
[17] 李峰,周广胜,曹铭昌.兴安落叶松地理分布对气候变化响应的模拟.应用生态学报,2006,17(12):2255-2260.
[18] 王娟,倪健.植物种分布的模拟研究进展.植物生态学报,2006,30(6):1040-1053.
[22] 马松梅,张明理,张宏祥,孟宏虎,陈曦.利用最大熵模型和规则集遗传算法模型预测孑遗植物裸果木的潜在地理分布及格局.植物生态学报,2010,34(11):1327-1335.
[23] 雷军成,徐海根.基于MaxEnt的加拿大一枝黄花在中国的潜在分布区预测.生态与农村环境学报,2010,26(2):137-141.
[24] 王运生,谢丙炎,万方浩,肖启明,戴良英.相似穿孔线虫在中国的适生区预测.中国农业科学,2007,40(11):2502-2506.
[28] 曹向锋,钱国良,胡白石,刘凤权.采用生态位模型预测黄顶菊在中国的潜在适生区.应用生态学报,2010,21(12):3063-3069.
[29] 杨波,薛跃规,唐小飞,王媛,张达敏.外来入侵植物飞机草在中国的适生区预测.植物保护,2009,35(4):70-73.
[30] 吴文浩,李明阳.基于生态位模型的松材线虫潜在生境预测方法研究.林业调查规划,2009,34(5):33-38.
[35] 刘宇,陈泮勤.张稳,胡非.一种地面气温的空间插值方法及其误差分析.大气科学,2006,30(1):146-152.
[37] 翁恩生,周广胜.用于全球变化研究的中国植物功能型划分.植物生态学报,2005,29(1):81-97.
[38] 张新时.植被的PE(可能蒸散)指标与植被-气候分类(二)——几种主要方法与PEP程序介绍.植物生态学与地植物学学报,1989,13(3):197-207.
[39] 倪健.中国亚热带常绿阔叶林优势种及常见种分布与Thornthwaite指标的关系.武汉植物学研究,1997,15(2):151-161.
[41] 邵慧,田佳倩,郭柯,孙建新.样本容量和物种特征对BIOCLIM模型模拟物种分布准确度的影响——以12个中国特有落叶栎树种为例.植物生态学报,2009,33(5):870-877.
[42] 许中旗,王义弘.蒙古栎研究进展.河北林果研究,2002,17(4):365-370.
[43] 郭建平,高素华,刘玲,周广胜.气候变化对蒙古栎生长和气候生产力的影响.资源科学,2005,27(5):168-172.
猜你喜欢
阅读与作文(小学高年级版)(2019年8期)2019-10-16
科学大众(中学)(2019年3期)2019-05-17
蒙古学问题与争论(2019年0期)2019-03-29
西域历史语言研究集刊(2018年0期)2018-11-09
汽车观察(2018年10期)2018-11-06
草原歌声(2017年1期)2017-04-23
科技知识动漫(2017年1期)2017-02-06
西南农业学报(2016年6期)2016-04-16
少儿科学周刊·少年版(2015年1期)2015-07-07
湖北农业科学(2014年21期)2014-12-22
