
长白山林线树种岳桦幼树叶功能型性状随海拔梯度的变化
2013-12-16 08:20:10胡启鹏郭志华孙玲玲
生态学报 2013年12期
胡启鹏,郭志华,孙玲玲,王 彬
(1.嘉汉林业(中国)投资有限公司,广州 510613;2.中国林业科学研究院湿地研究所,北京 100091;3.广东省生态环境与土壤研究所,广州 510650;4.亚热带森林培育国家重点实验室培育基地,临安 311300)
海拔是影响植物生理、结构和功能、代谢等重要的生态因子之一[1],随海拔高度变化,水热、光照、CO2分压等亦表现出梯度性变化,植物叶面积、叶氮(Nmass和Narea)、叶绿素含量、光合等叶特征表现出不同的适应性响应[2-6]。高海拔限制森林的分布,从而形成各种森林分布的界限,即高山林线[7],由于地理位置的特殊,其生态生理机制一直是探索的热点,如光合和呼吸作用、适应性、多样性、生长、解剖学等[8-11],而从叶功能型性状方面研究较少[12-15],特别在全球大气变化的影响下,林线树种在形态或生理上的响应则更为敏感[16],研究林线植物叶功能型性状对不同海拔梯度的响应,对揭示林线树种适应机制和分布原因具有重要生态学意义。
长白山森林生态系统是植被垂直梯度分布最典型之一,而亚高山岳桦林带是从森林过渡到苔原的一个过渡带,岳桦即为林线树种,以前的学者已经从种群[17-18]、群落[19]、分布[20]、林线变化[16,21]、生物量[22]等角度进行了研究,而叶特征方面主要从碳素供应[13]、非结构性碳水化合物[14]及生理[7]方面进行了报道。本文从不同海拔梯度上岳桦幼树叶功能型性状及可塑性方面研究出发,以探讨岳桦对海拔的响应和适应机制。
1 材料与方法
1.1 试验地概况
试验地位于长白山北坡亚高山岳桦林带,地处火山锥体中部,基岩有粗面岩、粗面角砾岩、流纹岩等,随着分布海拔的升高,常形成纯林,最后于苔原带接壤。沿海拔梯度,与岳桦伴生的灌木以牛皮杜鹃(Rhododendron aureum)和樾桔(V.vitis-idaea)为主等。草本植物比较发达,覆盖度70%—100%,多为耐寒植物。
岳桦林的土壤也属于一个过渡带,在与针叶树混交的林分带属于山地棕色针叶林土,随海拔升高,在1800m以上,土壤逐渐变为山地生草森林土,最后过渡到无林的山地苔原土。
1.2 试验地设置
根据长白山岳桦的分布特征,并参考邹春静等[20]和于大炮等[23]有关岳桦过渡带位置的研究,选取沿长白山北坡岳桦分布连续的7个采样点采样(表1),采样点布设在不同海拔处距林缘10—30m内,采样范围10—30m2。
1.3 试验材料
实验材料为岳桦(Betula ermanii)4—5年生幼树。幼树年龄通过查数轮生枝法获得。
岳桦,桦木科(Betulaceae)桦木属(Betula L.),落叶乔木。
1.4 试验方法
叶面积LA测定采用图纸法,具体方法:在待测植株上,每株采取中上部当年生叶片,在A4纸上画出叶片轮廓,然后沿轮廓裁剪并称重Cm,然后利用整张纸的面积ZL与质量Zm换算叶片面积。比叶重LMA=叶片干重/叶面积。用凯氏定氮法测叶片总氮含量。叶绿素含量、叶片脯氨酸、丙二醛(MDA)、抗坏血酸氧化物酶(ASA-POD)活性(APX)测定参照邹琦[24]。可溶性糖测定采用蒽酮比色法[25]。
可塑性指数(PI)等于岳桦叶功能型指标在海拔梯度上的最大值减去最小值再除以最大值[26]。
以上性状测定的样品均采自原生环境的4—5a幼树上,每个地点测定5株,取样时按不同方向上每株取10片当年生成熟、无病虫害叶,混匀,除LA和LMA外,其他均由液氮固定后带回实验室进行测定。取样时间在2007年8月3—5日。
各变量均以加减一个标准误表示,差异性水平(P<0.05)分析均采用SPSS 13.0。
2 结果
2.1 随海拔梯度升高叶面积与比叶重的变化
叶片作为植物接收光能进行碳同化的最主要器官,其面积的大小和薄厚是衡量植物功能性状的主要参数[2]。图1显示,随海拔升高,LA呈逐渐降低趋势,A5处下降较快,而LMA呈逐渐升高变化,其值在A7处达最高水平,与A1处最小值相比增加了53%。经检验LA和LMA差异均达到显著水平(P<0.05)。

图1 与岳桦叶面积及比叶重随海拔高度变化Fig.1 Responses of leaf area(LA)and leaf dry mass per area(LMA)of B.ermanii to different altitudes
2.2 叶绿素与叶氮
叶绿素是植物进行光合作用的载体,其含量大小影响植物的光合能力和光合器官的保护等[5]。表2显示,随海拔升高,叶绿素a(Chla)、叶绿素b(Chlb)及总叶绿素含量(Chl)均呈逐渐降低趋势,A7处达到最低,与A1处相比分别降低了45%、52%和47%,而类胡萝卜素Car和Car/Chl比值则呈上升趋势,说明Chl分配给Car的量增加,经检验叶绿素含量在不同海拔间差异均达到显著水平(P<0.05)。叶氮含量(Nmass和Narea)随海拔变化差异显著(P<0.05),均在A7处达最高水平,中部海拔相对偏低。分配到叶绿素中的N(Chl/Nmass)随海拔升高而降低,A7处较A1处下降47%,且差异达到显著水平(P<0.05)。
2.3 丙二醛、脯氨酸、抗坏血酸酶活性及可溶性糖
图2显示,随海拔高度升高,丙二醛(MDA)逐步升高,且差异显著(P<0.05),说明岳桦遭受到了高海拔逆境的限制,包括低温和紫外线辐射,从而导致活性氧增加。同时抗氧化物质脯氨酸Pro、可溶性糖DS、及抗坏血酸活性APX均随海拔升高明显增加,且显著(P<0.05),表明随海拔升高,岳桦受到胁迫越强的同时抗逆性也增强。

表2 岳桦叶绿素、类胡萝卜素及叶氮含量随海拔梯度的变化Table 2 Responses of chlorophyll,carotenoid and leaf nitrogen of B.ermanii to different altitudes

图2 岳桦叶片MDA、DS、APX、Rro随海拔高度变化Fig.2 Response of MDA、DS、APX、Rro of B.ermanii to different altitudes
2.4 可塑性比较
可塑性是指植物在适应过程中通过改变形态、生理等性状满足其表型与其生存环境相一致[27],一般高的可塑性与植物对环境具有高的潜在适应能力紧密相关[28]。表3显示,综合可塑性指数从大到小依次为:基于抗氧化物质(0.79)>基于叶绿素含量(0.49)>基于叶形态特征(0.47)>基于叶氮含量(0.39)。单项指标中,APX、Pro、DS和Car/Chl可塑性指数均高于0.6以上,Chlb、Nmass和 LMA 可塑性指数在0.6—0.5之间,其余均低于0.5,说明随海拔升高,岳桦向上的发展更倾向于自身的保护,生长次之。
3 讨论
叶片是植物捕获光能,进行光合作用的主要场所,LA和LMA作为植物碳收获策略的关键叶特征之一,其大小直接与光合碳同化和适应相关。研究表明[26,29],较小的叶面积和叶片较厚的植物,光合能力一般较低,因为较小的LA降低了光能的吸收和消耗,但可以抵御或避免恶劣环境的胁迫。沿长白山海拔升高,岳桦LA逐渐变小,LMA逐渐增大,意味着叶片越来越厚,这样可以减少高海拔低温、紫外线等不利环境因子的影响,降低能的消耗,以保证自身的正常生存,是一种适应的表现,这与多数植物随海拔高度变化一致[2,5,27,30],但叶形态中LMA较大可塑性指数表明,岳桦适应不同海拔环境更借助于叶片薄厚程度来捕获或抵消光能。
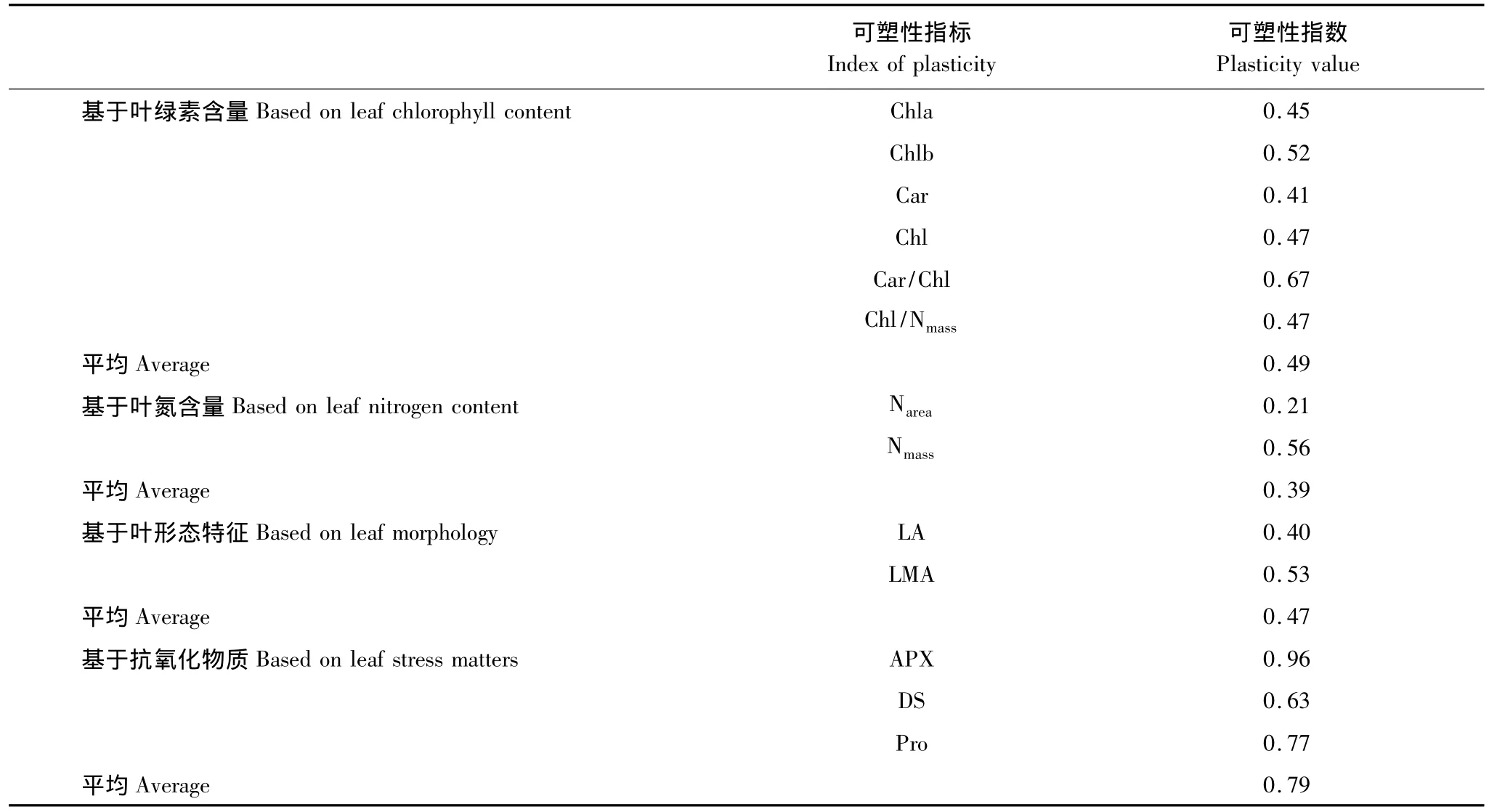
表3 岳桦叶片形态指标可塑性指数比较Table 3 Comparison of leaf functional characteristics plasticity index of B.ermanii
叶绿素含量的高低直接影响植物光合作用的表现。已有研究发现,植物叶绿素含量(a、b、a+b)沿海拔梯度升高呈下降趋势[31],这与本研究结果一致,亦印证了吴栋栋等[7]对岳桦叶绿素随海拔变化的研究结果。因为随海拔升高,辐射增强,叶绿素降低可以减少对光能的吸收,减弱对光合系统的压力,从而避免潜在的光抑制和过氧化[32]。Car具有吸收光能和保护叶绿素分子免遭光氧化损伤的功能[33],随海拔升高岳桦Car增加,但其较低的可塑性指数意味着其吸收光能用于光合作用的功能较低,主要是吸收多余的光能,保护光合器官,这从Car/Chl较高的可塑性指数可以得到印证,说明光合作用正常进行的前提是保护光合器官免受损。而Chlb较高的可塑性指数,表明岳桦在光合适应中更依赖吸收蓝紫光的Chlb吸收和传递光能,其次是Chla。叶氮与植物的光合作用密切相关,在不同光合器官组分分配中具有非常重要的作用[34]。研究表明Nmass的多寡直接决定着叶片光合能力的高低[35-36],主要原因是氮素在光合器官中占有较大比重。而Narea与冠层位置以及光子通量密度密切相关,具体表现在适应不同环境下的荫生叶或阳生叶[37]。但与Narea相比,岳桦较高的Nmass可塑性指数意味着其在随海拔适应过程中更依赖于Nmass对光合器官中的贡献。岳桦在高海拔表现出较高的Narea和Nmass,但分配到叶绿素的量(Chl/Nmass)却降低,,说明高海拔充足的光照使得岳桦不必在光合器官中投资太多的氮,而把部分氮投资于保护构造上以防止高温或强光损伤和失水过多[38],而Chl/Nmass和Chl相对较低的可塑性指数亦说明岳桦在向上适应的过程中,并不是为了积累光能而生长,而是为了减小光合器官的损伤而生存。在低海拔,胁迫相对降低,植物将更多地氮投资到了光合器官中,以便尽可能地吸收光照,维持光合生产[39],这与低海拔较低的LMA和Car一致。而海拔1800—1900m间Narea和Nmass含量最低,可能与岳桦的起源有关,野外调查亦发现,此海拔区间也是岳桦分布最多,最密集的区域。
MDA是膜质过氧化产物主要产物之一,表征植物对逆境的反映强弱。在植物遭受逆境的过程中,其量会增加,破坏蛋白质和酶的结构以及能引起叶绿体和过氧化物体中一些酶的活性下降,从而抑制光合作用[40-41]。随海拔梯度上升,岳桦叶片MDA逐渐增大,表明岳桦向上发展的过程中遭受到了高海拔逆境的限制,包括低温和紫外线的胁迫,这些导致活性氧增加,光合能力可能降低,这与吴栋栋等[7]对高海拔岳桦研究结果类似,但与其在低海拔结果并不一致,他认为岳桦在低海拔较高的活性氧是高温高湿刺激所致。但野外观测发现,在海拔1500m以下,特别在路边及开阔的环境中,岳桦的发展较快,说明岳桦是喜光树种,而温度被认为是海拔梯度上限制植物分布和生长的主要因子[42],因此可以认为低海拔高温对岳桦胁迫较小或没有,反而是其向下发展的有利因子。虽然逆境能引起植物活性氧增加,代谢紊乱,但相应地植物体内会形成抗氧化系统,保护植物免受活性氧危害[43]。且活性氧的清除由非酶促系统和酶促清除系统协同完成[41]。随海拔升高,低温等环境越来越恶劣,可溶性糖(DS)和脯氨酸(Pro)主动积累,这样可以降低渗透势和冰点[44],提高岳桦对低温逆境的抗性,抗坏血酸酶(APX)活性随海拔升高而增强则可以消除活性氧对岳桦生长的最大危害,最大限度保护植物正常生长,这与Car变化一致,但在高海拔1900m以上,APX活性变化幅度较小,差异并不显著,说明在高海拔地区,随着环境不断恶化,生理胁迫程度可能会逐渐超过了其内部保护系统(主要为酶促系统)的承受和抵御能力,不利于岳桦的生长发育,阻碍其继续向更高海拔分布,这也可能导致林线的形成。而APX活性最大的可塑性指数表明,岳桦对低温等环境的适应更依赖于酶促清除系统,而非酶促系统(DS和Pro)的变化则更能体现出岳桦对恶劣环境的一种反应。同时岳桦叶片各项平均可塑性指数指标中,抗氧化物质的可塑性指数最大,叶绿素和叶形态次之,表明随海拔升高,岳桦林以保护自身的生存为最主要的策略机制,然后以吸收光能的Chlb及LMA指标为主要生长策略。
总之,随长白山海拔升高,岳桦受到的环境胁迫逐渐加重,在叶功能型状方面形成了一定的适应和响应机制,表现为随海拔升高,叶片LMA增加、叶面积逐渐变小、Chl(a、b、a+b)降低和Car、Car/Chl和Chl/Nmass升高,Narea和Nmass在海拔1800—1900m间最低,在高海拔将更多的氮投资于保护光合器官,而低海拔则更倾向于光合生产。抗氧化物质DS、Pro和APX活性随海拔升高而增加,负责对活性氧的抵御和清除,但高海拔地区(1900m以上),APX活性变化较小,而MDA显著增加,意味着随海拔升高,恶劣环境对岳桦的胁迫可能逐渐超过体内的保护系统,缓冲岳桦继续向上生长和分布,这也可能是岳桦林线形成的原因之一。同时可塑性指数表明岳桦的向上分布的过程中,其适应环境更依赖抗氧化物质,即生存为主,生长次之。
[1] Pigott C D.Experimental studies on the influence of climate on the geographical distribution of plants.Weather,1975,30(3):82-90.
[2] Hölscher D,Schmitt S,Kupfer K.Growth and leaf traits of four broad-leaved tree species along a hillside gradient.Forstwissenschaftliches Centralblatt,2002,121(5):229-239.
[3] Hikesaka K,Nagamatsu D,Ishii H S,Hirose T.Photosynthesis-nitrogen relationships in species at different altitudes on Mount Kinabalu,Malaysia.Ecological Research,2002,17(3):305-313.
[4] Crain J M,Lee W G.Covariation in leaf and root traits for native and non-native grasses along an altitudinal gradient in New Zealand.Oecologia,2003,134(4):471-478.
[5] Qi J,Ma K M,Zhang Y X.The altitudinal variation of leaf traits of Quercus liaotungensis and associated environmental explanations.Acta Ecologica Sinica,2007,27(3):930-937.
[6] Llorens L,Peňuelas J,Beier C,Emmett B,Estiarte M,Tietema A.Effects of an experimental increase of temperature and drought on the photosynthetic performance of two ericaceous shrub species along a North-South European gradient:climate change impacts on terrestrial ecosytems(CLIMOOR).Ecosystems,2004,7(6):613-624.
[7] Wu D D,Zhou Y B,Yu D P,Dai G H.Physiological response of Betula ermanii at different altitudes in Changbai Mountain.Acta Ecologica Sinica,2009,29(5):2279-2285.
[8] Billings W D.Arctic and alpine vegetation:Plant adaptation to cold summer climates//Ives J D,Barry R G,eds.Arctic and Alpine Environments.London:Methuen,1974:403-443.
[9] Myers N,Mittenneier R A,Mittenneier C G,da Fonseca G A B,Kent J.Biodiversity hotspots for conservation priorities.Nature,2000,403(6772):853-858.
[10] Billings W D,Mooney H A.The ecology of arctic and alpine plants.Biological Reviews,1968,43(4):481-529.
[11] Woodward F I.Ecophysiological studies on the shrub Vaccinium myrtillus L.taken from a wide altitudinal range.Oecologia,1986,70(4):580-586.
[12] Li F l,Bao W K,Liu J H,Wu N.Eco-anatomical characteristics of Sophora davidii leaves along an elevation gradient in upper Minjiang River dry valley.Chinese Journal of Applied Ecology,2006,17(1):5-10.
[13] Zhou Y B,Wu D D,Yu D P.Carbon Supply Status in the Betula ermanii in Changbai Mountain.Scientia Silvae Sinicae,2010,46(3):161-165.
[14] Zhou Y B,Wu D D,Yu D P,Sui C Y.Variations of nonstructural carbohydrate content in betula ermanji at different elevations of Changbai Mountain,China.Chinese Journal of Plant Ecology,2009,33(1)118-124.
[15] Luo T X,Luo J,Pan Y D.Leaf traits and associated ecosystem characteristics across subtropical and timberline forests in the Gongga Mountains,Eastern Tibetan Plateau.Oecologia,2005,142(2):261-273.
[16] Zhou X F,Wang X C,Han S J,Zou C J.The effect of global climate change on the dynamics of Betula ermanii-Tundra ecotone in the Changbai Mountains.Earth Science Frontiers,2002,9(1):227-231.
[17] Zou C J,Han S J,Zhou Y M,Wang X C,Chen Y L.Study on ecological characteristics of Betula ermanii population in ecotone.Chinese Journal of Applied and Environmental Biology,2001,7(1):1-6.
[18] Wang X C,Han S J,Zou C J,Zhou X F.Geostatistical analysis of the pattern of Betula ermanii population in Changbai Mountain.Chinese Journal of Applied Ecology,2002,13(7):781-784.
[19] Wang Z,Xu Z B,Li X.The main forest types and one of its community structure characteristics in the north slope of Changbai Mountain.For Ecosyst Res,1980:25-39.
[20] Zou C J,Wang X C,Han S J.Position of Betula ermanii population ecotone in Changbai Mountains.Chinese Journal of Applied Ecology,2004,15(12):2217-2220.
[21] Wang X C,Zhou X F,Sun Z H.Research advances in the relationship between alpine timberline and climate change.Chinese Journal of Ecology,2005,24(3):301-305.
[22] Deng K M,Shi P L,Yang Z L.Biomass allocation and net primary productivities at Treeline Ecotone on the Changbai Mountains,Northeast China.Journal of Natural Resources,2006,21(6):942-948.
[23] Yu D P,Zhou L,Dong B L,Dai L M,Wang Q L.Structure and dynamics of Betula ermanii population on the Northern slope of Changbai Mountain.Chinese Journal of Ecology,2004,23(5):30-34.
[24] Zou Q.Experimental Guidance of Plant Physiology and Biochemistry.Beijing:China Agriculture Press,1995:30-35.
[25] Li H S.Principles and Techniques of Plant Physiological Biochemical Experiment.Beijing:Higher Education Press,2000:1-30.
[26] Hu Q P,Guo Z H,Li C Y,Ma L Y.Review on phenotypic plasticity in Plants response to abiotic factors.Scientia Silvae Sinicae,2008,44(5):135-142.
[27] Schmitt J,Stinchcombe J R,Heschel M S,Hube H.The adaptive evolution of plasticity:phytochrome-mediated shade avoidance responses.Integrative and Comparative Biology,2003,43(3):459-469.
[28] Valladares F,Wright S J,Lasso E,Kitajima K,Pearcy R W.Plastic phenotypic response to light of 16 congeneric shrubs from a Panamanian rainforest.Ecology,2000,81(7):1925-1936.
[29] Poorter L,Bongers F.Leaf traits are good predictors of plant performance across 53 rain forest species.Ecology,2006,87(7):1733-1743.
[30] Bradshaw A D.Unravelling phenotypic plasticity-why should we bother?New Phytologist,2006,170(4):644-648.
[31] Zhou D W,Zhu W Y,Teng Z H,Shi S B,Liu J Q,Han F.Antioxidative compounds of Polygonum viviparum L.from different altitudes.Chinese Journal of Applied and Environmental Biology,2003,9(5):489-492.
[32] Osone Y,Tateno M.Nitrogen absorption by roots as a cause of interspecific variations in leaf nitrogen concentration and photosynthetic capacity.Functional Ecology,2005,19(3):460-470.
[33] Liu Q,Wang Q C,Xu J,Sun J.Effects of exogenous spermidine and spermine on Celastrus orbiculatus antioxidant system under soil NaHCO3stress.Chinese Journal of Applied Ecology,2009,20(3):549-554.
[34] Kitao M,Lei TT,Koike T,Tobita H,Maruyama Y.Susceptibility to photoinhibition of three deciduous broadleaf tree species with different successional traits raised under various light regimes.Plant,Cell and Environment,2000,23(1):81-89.
[35] Körner C.The nutritional status of plants from high altitudes.Oecologia,1989,81(3):379-391.
[36] Reich P B,Walters M B,Ellsworth D S.Leaf life-span in relation to leaf,plant,and stand characteristics among diverse ecosystems.Ecological Monographs,1992,62(3):365-392.
[37] Anten N P R,Hirose T.Limitations on photosynthesis of competing individuals in stands and the consequences for canopy structure.Oecologia,2001,129(2):186-196.
[38] Zhang L,Luo T X.Advances in ecological studies on leaf lifespan and associated leaf traits.Acta Phytoecologica Sinica,2004,28(6):844-852.
[39] Ackerly D D,Knight C A,Weiss S B,Barton K,Starmer K P.Leaf size,specific leaf area and microhabitat distribution of chaparral woody plants:contrasting patterns in species level and community level analyses.Oecologia,2002,130(3):449-457.
[40] Dhindsa R S,Plumb-Dhindsa P L,Reid D M.Leaf senescence and lipid peroxidation:effects of some phytohormones,and scavengers of free radicals and singlet oxygen.Physiologia Plantarum,1982,56(4):456-457.
[41] Wang Y P,Huang H Y,An L Z,An L Z,Wang D,Zhang F.Antioxidative responses to different altitudes in leaves of alpine plant Polygonum viviparum in summer.Acta Physiologiae Plantarum,2009,31(4):839-848.
[42] Angert A L.Growth and leaf physiology of monkeyflowers with different altitude ranges.Oecologia,2006,148(2):183-194.
[43] Salin M L.Toxic oxygen species and protective systems of the chloroplast.Physiologia Plantarum,1988,72(3):681-689.
[44] Song G S,Sun Z F,Sun L,Du K M,Wang X.Comparison between physiological properties and cold tolerance under low temperature treatment during different growing stages of rice in northeast central region of China.Acta Ecologica Sinica,2011,31(13):3788-3795.
参考文献:
[5] 祁建,马克明,张育新.辽东栎(Quercus liaotungensis)叶特性沿海拔梯度的变化及其环境解释.生态学报,2007,27(3):930-937.
[7] 吴栋栋,周永斌,于大炮,戴冠华.不同海拔长白山岳桦的生理变化.生态学报,2009,29(5):2279-2285.
[12] 李芳兰,包维楷,刘俊华,吴宁.岷江上游干旱河谷海拔梯度上白刺花叶片生态解剖特征研究.应用生态学报,2006,17(1):5-10.
[13] 周永斌,吴栋栋,于大炮.长白山岳桦体内碳素供应状况.林业科学,2010,46(3):161-165.
[14] 周永斌,吴栋栋,于大炮,隋琛莹.长白山不同海拔岳桦非结构碳水化合物含量的变化.植物生态学报,2009,33(1):118-124.
[16] 周晓峰,王晓春,韩士杰,邹春静.长白山岳桦苔原过渡带动态与气候变化.地学前缘,2002,9(1):227-231.
[17] 邹春静,韩士杰,周玉梅,王晓春,陈永亮.过渡带中岳桦种群生态特征的研究.应用与环境生物学报,2001,7(1):1-6.
[18] 王晓春,韩士杰,邹春静,周晓峰.长白山岳桦种群格局的地统计学分析.应用生态学报,2002,13(7):781-784.
[19] 王战,徐振邦,李昕.长白山北坡主要森林类型及其群落结构特点之一.森林生态系统研究,1980,25-39.
[20] 邹春静,王晓春,韩士杰.长白山岳桦种群过渡带位置的研究.应用生态学报,2004,15(12):2217-2220.
[21] 王晓春,周晓峰,孙志虎.高山林线与气候变化关系研究进展.生态学杂志,2005,24(3):301-305.
[22] 邓坤枚,石培礼,杨振林.长白山树线交错带的生物量分配和净生产力.自然资源学报,2006,21(6):942-948.
[23] 于大炮,周莉,董百丽,代力民,王庆礼.长白山北坡岳桦种群结构及动态分析.生态学杂志,2004,23(5):30-34.
[24] 邹琦.植物生理学与生物化学实验指导.北京:中国农业出版社,1995:30-35.
[25] 李合生.植物生理生化实验原理和技术.北京:高等教育出版社,2000:1-30.
[26] 胡启鹏,郭志华,李春燕,马履一.植物表型可塑性对非生物环境因子的响应研究进展.林业科学,2008,44(5):135-142.
[31] 周党卫,朱文琰,滕中华,师生波,刘健全,韩发.不同海拔珠芽蓼抗氧化系统的研究.应用与环境生物学报,2003,9(5):489-492.
[33] 刘强,王庆成,徐静,孙晶.外源亚精胺和精胺对NaHCO3胁迫下南蛇藤抗氧化系统的影响.应用生态学报,2009,20(3):549-554.
[38] 张林,罗天祥.植物叶寿命及其相关叶性状的生态学研究进展.植物生态学报,2004,28(6):844-852.
[44] 宋广树,孙忠富,孙蕾,杜克明,王夏.东北中部地区水稻不同生育时期低温处理下生理变化及耐冷性比较.生态学报,2011,31(13):3788-379.
猜你喜欢
昆明医科大学学报(2022年1期)2022-02-28 07:43:34
散文诗(2021年22期)2022-01-12 06:14:04
昆明医科大学学报(2021年8期)2021-08-13 08:59:20
昆明医科大学学报(2021年6期)2021-07-31 07:40:08
学生天地(2020年34期)2020-06-09 05:50:38
辽金历史与考古(2019年0期)2020-01-06 07:44:54
环球时报(2019-05-23)2019-05-23 06:17:36
少儿美术(快乐历史地理)(2018年7期)2018-11-16 05:31:12
西南医科大学学报(2015年1期)2015-08-22 13:01:48
中国工程咨询(2015年8期)2015-02-16 06:39:04
