
上海人工林林下木本植物更新格局及其影响因素
2013-12-02 02:29郭雪艳达良俊惠晓萍
华东师范大学学报(自然科学版) 2013年2期
张 净,郭雪艳,达良俊-3,王 婕,惠晓萍,耿 晔
(1.华东师范大学 环境科学系,上海 200062;2.华东师范大学 上海市城市化生态过程与生态恢复重点实验室,上海 200062;3.华东师范大学 天童森林生态系统国家野外科学观测研究站,上海 200062;4.上海市林业总站,上海 200072)
0 引 言
上海作为我国森林资源最贫乏的城市之一,进入21世纪以来,随着生态型城市的建设,城市绿化建设取得突破性进展.截止2011年底,全市森林覆盖率达到12.6%(上海市统计局,2012),初步构建了森林生态系统框架.但在上海市人工林建设之初,由于受人为因素的影响,多表现为群落类型单一、结构简单、物种贫乏,形成了大量苗圃式、纯林式人工植被[1].因此,通过林相改造与人工林自然恢复相结合,实现功能优化,已势在必行.
森林更新作为重要的生态学过程,一直是森林生态系统研究中的主要领域之一[2-6].随着城市人工林的建成与发展,一些依靠重力、风力及动物散布种子的木本植物实生幼苗开始自然更新,人工林逐步进入自然恢复演替阶段[1],其影响着群落的物种组成[7-9]、结构[10]和动态变化[11-13],是种群得以增殖、扩散、延续和维持群落稳定的一个重要生态过程[14].本文研究上海城市人工林更新层种类组成、多样性特征及其生境特点,并探讨其更新格局成因,借鉴潜在自然植被,为进一步制定林相改造计划,建设高生物量、高多样性、高生态效益和少管护的城市森林提供理论依据和技术支撑.
1 研究区域概况及研究方法
1.1 研究区域概况
上海市(N30°40′—31°53′,E120°51′—122°12′)位于暖温带向亚热带的过渡带,属于亚热带北缘季风气候区,降水丰沛,温和湿润,具有显著的海洋性气候特征.地带性植被为含有落叶成分的常绿阔叶林,但在长期人为活动影响下,其仅在大金山岛及佘山地区有部分分布.人工植被成为上海市的主要植被类型,其物种组成较单一,群落类型多样但结构简单,并在高强度的人为干扰及管护下维持着特有的景观和分布格局[15-17].
1.2 林分调查
于2007—2009年以上海市中心城区和外环绿带人工林为对象,选取典型、均质样地,其中上海市中心城区478个样方,参照方和俊[17]等对上海城市绿地群落的研究,选择中心城区人工绿地48个,包含24个公园绿地、3个防护绿地、7个附属绿地,14个其他绿地,共4个类型;外环绿带430个样方,包含乔木样方386个,灌木样方44个,共计调查样方908个,样方面积的设置乔木林为10m×10m和20m×20m,灌丛设置为5m×5m和10m×10 m.调查采用法瑞学派调查法和每木调查法相结合的方法,根据群落垂直结构目测分层,记录林木层各层中植物以及更新层木本植物实生幼苗的种类、最大高度、盖度.
使用W.E.T.土壤三参数速测仪测定群落内的土壤湿度,测量深度为10cm厚土壤层;同时使用光量子仪测定群落内光照和群落外裸地光照值,为减少生境因子的测定误差,测定均在至少连续一周无雨时进行,各样地均重复5次.
1.3 数据分析方法
物种多样性指数采用物种丰富度(S),均匀度指数(E)、Shannon-Wiener指数(H)以及Simpson指数(D),以上指数在PC-ORD(Version 5.0)软件包上完成.基于物种相对优势度,运用PC-ORD软件,采用典范对应分析法(CCA)分析更新层木本植物分布与环境因子间相关性,所有差异显著性检验均在SPSS 18.0软件上完成.
为进行不同取样面积间物种总数的对比分析,对物种总数进行标准化处理,处理公式为:
N=N1/ln n,
式中:N1为样方中出现的物种总数;N为标准化的物种总数;n为样方数(下同).
2 结 果
2.1 更新层与林木层物种丰富度及其出现频率
2.1.1 更新层与林木层种类比较
本次共调查908个样方,共记录林木层木本植物有60科107属149种(含变型、变种和栽培变种,下同),所记录的植物中自然分布的乡土树种78种,占总种数的52.35%,其中上海地区自然分布的有37种,周边江浙地区的有41种;园艺栽培种的有71种.
共记录更新层木本植物有55科87属101种,其中隶属上海地区自然分布的乡土树种有36种,周边江浙地区的有28种,共计64种,占总种数的63.37%;园艺栽培起源的种类有37种,占总种数的36.63%(附表详见电子版,http://xblk.ecnu.edu.cn).
分析更新层与林木层植物的关系(见表1),林木层中有97种植物可进行自我更新,占林木层总种数的65.10%,其中可自我更新的落叶针叶乔木、常绿针叶乔木、落叶阔叶乔木、常绿阔叶乔木、落叶灌木和常绿灌木分别占到了林木层各自生活型总数的33.33%、33.33%、83.33%、69.23%、43.75%和60.00%;此外,更新层有4种植物在所调查样方林木层中无母株分布,分别为大青(Clerodendrum cyrtophyllum)、柘树(Cudrania tricuspidata)、豪猪刺(Berberis julianae)和野花椒(Zanthoxylum simullans),仅占人工林树种总数的2.68%.
2.1.2 更新层主要物种出现频率
大部分的物种为出现频率不足1%的偶见种,出现频率大于1%的常见种为36种,仅占35.6%.除9个园艺栽培种外,上海及周边江浙地区自然分布的乡土树种为27种,占常见种的75.0%,其中常绿阔叶乔木女贞和香樟在一半以上的群落中出现,频率高达58.6%和53.7%;落叶阔叶乔木出现种类最多达14种,朴树和构树出现频率大于30%,桑树、栾树、乌桕和苦楝为5%~20%,合欢、无患子和臭椿等8个物种均在5%以下;灌木树种中,海桐为10.9%,小蜡为6.6%,蚊母、胡颓子和火棘等其余8个种均小于5%;常绿针叶乔木罗汉松为3.6%.园艺栽培种中,常绿乔木棕榈的出现频率最高,为29.6%;落叶阔叶乔木榆树、刺槐和鸡爪槭分别为6.1%、3.1%和2.2%;其余5种均为常绿阔叶灌木,其中八角金盘为13.2%,桂花、珊瑚树和红叶石楠等4种常绿灌木均小于3%(见表2).

表1 更新层与林木层植物种类比较Tab.1 Comparison of woody plants between regeneration layer and tree layer
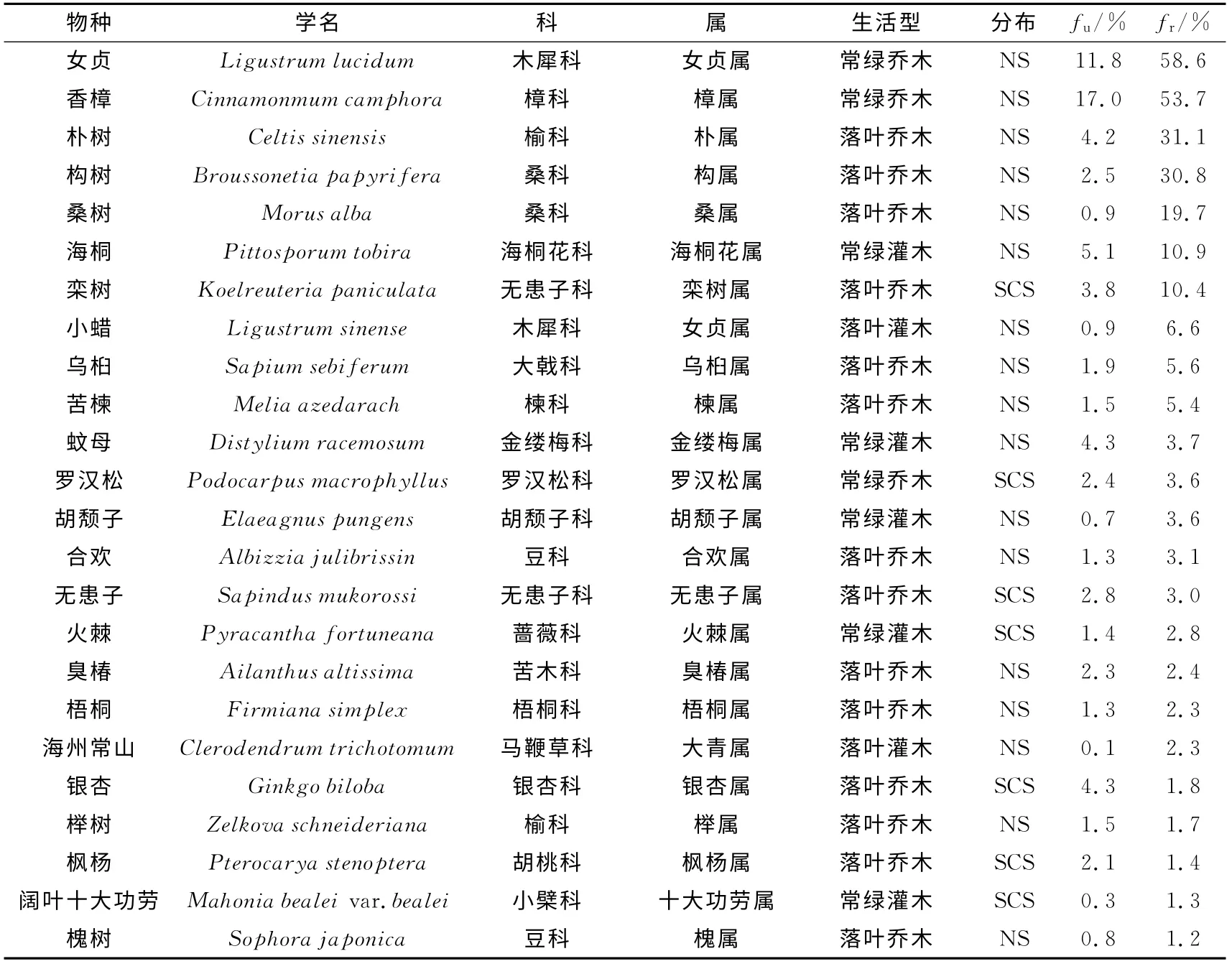
表2 更新层木本植物主要树种及其出现频率(f≥1%)Tab.2 Main species and their occurrence frequency of seedlings in understory regeneration layer(f≥1%)

续表2
2.2 更新层物种多样性特征
在908个样方中,有99个更新层中无木本植物分布,占10.9%;29.5%的样方更新层组成物种数为1~2种,而林木层高达70.6%;更新层树种数为3~4、5~7和8种以上的样方数均高于林木层,样方更新层较林木层物种丰富度更高(见图1).
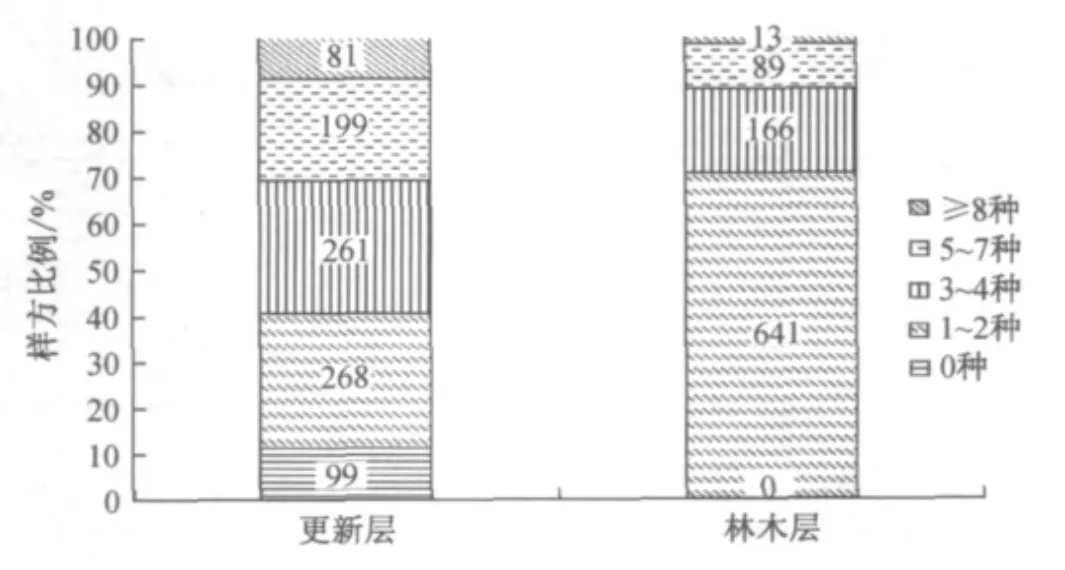
图1 更新层及林木层群落丰富度Fig.1 Community richness of upper layer and regeneration layer
2.2.1 不同群落类型更新层物种多样性特征
以建群种生活型为依据,在群落外貌水平上可划分为9个植被型组[18].落叶阔叶林(DBF)和常绿阔叶林(EBF)比例最高,为别为26.4%、24.7%;常绿针叶林(ECF)、落叶针叶林(DCF)、针阔混交林(BCMF)、竹林(BF)比例较少,为4.0%~6.0%,而作为地带性植被的常绿落叶阔叶混交林(EDBF)的比例并不高,仅为10.0%;群落组成结构比较单一的常绿灌丛(ES)、落叶灌丛(DS)比例也较低,都在10%以下.
9种不同类型群落中,常绿落叶阔叶混交林和落叶阔叶林更新层物种总数最多,分别高达12.86和12.41;针阔混交林和常绿阔叶林次之,分别为10.91和10.53;其余群落类型均小于10.不同类型群落的物种丰富度均高于林木层,其中林木层丰富度显著高于其它群落类型的针阔混交林和常绿落叶阔叶混交林,其更新层物种多样性也要显著高于其它群落类型(p<0.05);其次为落叶阔叶林和常绿阔叶林;林木层物种单一的竹林各多样性指数仅次于落叶阔叶林和常绿阔叶林;落叶针叶林各多样性指数略高于常绿针叶林,均低于竹林;落叶灌丛和常绿灌丛多样性指数要显著低于其它群落类型(见表3).

表3 不同群落类型人工林更新层木本植物的物种多样性Tab.3 Species diversity of understory woody plants between different communities
2.2.2 不同结构类型更新层物种多样性特征
在群落垂直结构上,可划分为乔—灌—草(T-S-H)结构、乔—草(T-H)结构、灌—草(SH)结构3个类型.其中,乔—草结构和乔—灌—草结构比例较高,分别为43.8%和33.9%;灌—草结构最少,所占样方比例为22.2%.
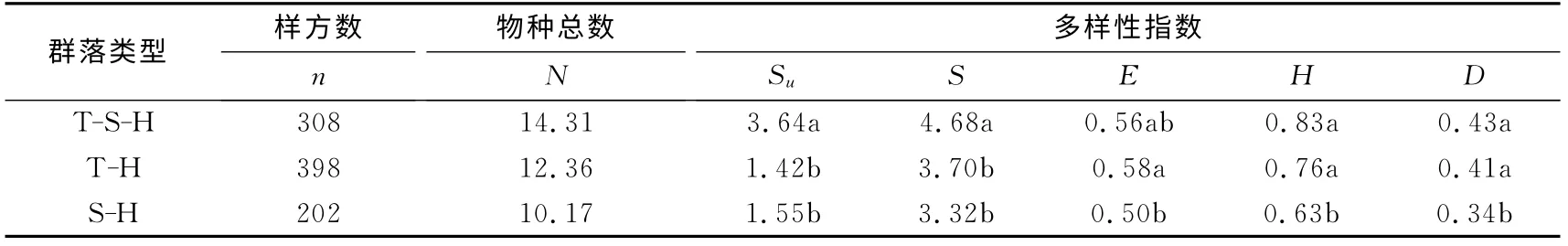
表4 不同结构类型人工林更新层木本植物的物种多样性Tab.4 Species diversity of understory woody plants between different community structure
不同结构类型更新层物种总数表现为乔—灌—草结构最高,为14.31,其次是乔—草结构的12.36,灌—草结构较少,为10.17.3种群落结构的更新层丰富度均高于林木层,表现为乔—灌—草结构>乔—草结构>灌—草结构,且乔—灌—草结构物种丰富度显著高于乔—草、灌—草结构(p<0.05).更新层的物种多样性指数变化趋势基本一致,Shannon-wiener指数(H)、Simpson指数(D)均表现为乔—灌—草结构>乔—草结构>灌—草结构,Pielou(E)指数则表现为乔—草结构>乔—灌—草结构>灌—草结构,其中乔—灌—草和乔—草结构的E、H、D指数均显著高于灌—草结构(p<0.05)(见表4).
2.3 更新层物种组成与生境因子相关性
对人工林林下生境因子进行显著性和相关性分析,所分析的生境因子中生物因素主要包括林木层盖度、灌木层盖度和草本层盖度;非生物因素为相对光照强度和土壤湿度.
相关性分析发现(见表5),光照与林木层盖度呈极显著负相关(p<0.01),林木层盖度越大,林内郁闭度越高,相对光照强度也越低;土壤湿度与草本层盖度呈极显著正相关(p<0.01),草本层盖度越大,土壤湿度越大,林内生境就越阴湿;林木层盖度与灌木层盖度呈极显著负相关(p<0.01),林木层郁闭度过高,光照不足,会限制林下灌木的生长;灌木层盖度与草本层盖度呈显著负相关.
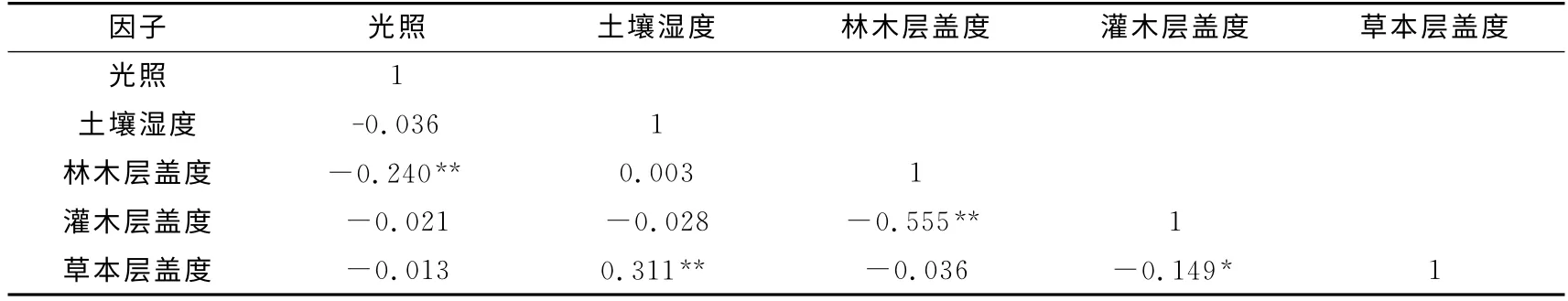
表5 生境因子相关分析Tab.5 Analysis of correlation between habitat factors

图2 不同群落类型自然更新层分布与生境因子相关性Fig.2 The relationship between the distribution of species under communities and habitat factors
在此基础上,基于更新层物种相对优势度与生境因子间的相关性典范对应分析(CCA),结果显示影响群落更新层物种组成的因子为光照强度、土壤湿度和草本层盖度,其中与光照强度的相关性最大,呈负相关关系,其次是土壤湿度和草本层盖度(见图2).
3 讨 论
3.1 人工林林下更新层物种多样性成因
林木层种类的65.1%在更新层中有分布,共97种,占更新层种类的96.0%,表明林木层植物对林下更新层的形成起着重要作用,是更新层的稳定种源,并使得种群得以自然更新,而林木层中无分布的大青、柘树、豪猪刺和野花椒等4个物种,由于在所调查样方周边有母株的分布,在更新层中也有出现,种源的存在与否不仅成为限制林下植被自然更新的主要因素,也决定了更新层种类组成[1,19].同时更新层多样性格局也受群落类型及群落结构的影响,常绿落叶阔叶混交林和针阔混交林上层物种丰富度最高,种源较丰富,因此更新层物种多样性也高于其它群落类型;而物种组成较单一的常绿灌丛和落叶灌丛,由于缺乏种源,从而限制了其林下更新.群落结构也是决定更新层物种多样性的另一重要因素,由于乔—灌—草结构较乔—草结构和灌—草结构上层物种更为丰富,因此其更新层物种多样性也较高.
城市人工林大多处于造林初期,呈片段化、斑块状或孤立分布,不利于种子的扩散,成为人工林近自然恢复过程中植物定居的限制因子[20],而物种的自身生物学特征如种子的散布方式也是决定其自然更新格局的重要因素[21].本研究中更新层木本植物的出现频率不尽相同,除香樟、女贞、朴树、构树、桑树、海桐、栾树、棕榈和八角金盘等9个物种出现频率大于10%,其余种出现频率多小于5%,且出现频率大于1%的物种也仅为36种,主要为在上海广泛种植有着丰富种源的树种如香樟、女贞等以及构树、桑树、朴树等依靠鸟类和风力散布种子的物种.而上海地区地带性顶级物种,如红楠(Machilus thunbergii)、苦槠(Castanopsis sclerophylla)、青冈(Cyclobalanopsis glauca)和冬青(Ilex purpurea)等物种在人工林中较少出现,这主要是由于片段化导致的种源、传播距离[22]和种子散布方式[23]等因素,限制了这些常绿阔叶树种在人工林自然恢复过程中的出现.
3.2 人工林自然更新的生境限制
种源和种子散布方式是限制物种定居的首要因素[4],同时外部环境条件也是重要的影响因子[19].种子萌发和幼苗的生长、发育、定居是植物生活史中对外界环境压力反应最为敏感的时期,是决定自然更新的重要阶段[24,25].适宜的光照条件是种子萌发和幼苗定居的关键,光照的变化能引起林地微环境湿度、温度变化,凡是影响光照条件的因子都会影响种子萌发和幼苗生长[26],城市人工林由于建设之初设计不合理,林分密度过高,其大多数林下郁闭度较高,且在某些群落内灌木成片密植,光照不足[27],限制了阳性物种的更新.树种的耐荫性不同,更新策略不同,喜光树种主要以灾变性的大型干扰更新方式更新,大多数顶级种以林窗更新方式更新,而一些耐荫树种以连续更新方式更新[28],香樟、女贞、栾树和海桐等耐荫性较好的植物能在林下快速定居,并迅速成为林下优势种;而随着林下郁闭度的不断增加,桑树、构树和朴树等阳性树种,由于没有充足的光照而个体数量逐渐减少.更新层木本植物出现频率较高的物种主要为常绿耐荫树种,可见林冠的存在对于出现耐荫树种的更新显然是有正面的作用,对于耐荫树种,上层林密,抑制了林下喜光物种的生长,减少了幼苗的竞争,因此有利于这些耐荫树种的天然更新[29].此外人工竹林多为散生,虽然其物种组成单一,但林下光照较充足,有利于周围物种的侵入定居,因此更新层物种多样性较高.
森林更新过程中土壤条件的异质性影响着种子的休眠、萌发与更新幼苗的发生格局[1].土壤湿度增大时,有利于种子萌发,并能提高幼苗成活率[11,30],上海市人工林群落自然更新与土壤湿度显著相关,种子散布进入林内后,能否成功定居很大程度上取决于林内生境条件,充足的水分条件有利于种子萌发和幼苗定居,从而促进更新;而更新层草本植物盖度越大,对土壤有限养分的竞争越激烈,间接影响了本本植物的萌发生长,因此林下草本植物盖度过大,在一定程度上会限制木本植物的更新.
3.3 建议
对上海来说,在今后人工林的总体规划建设中,以“零管护”和“少管护”为目标,划出植被发育较好的斑块进行保护,通过绿色廊道连接各个组团,提高群落之间的镶嵌性,同时增加人工植被的群落丰富度,优化城市植被的群落结构,从而形成完整的城市绿地网络系统,减少城市生境片断化导致的种源不足及种子难以散布等因素对植被恢复的限制[1].在植被恢复过程中,应遵循自然演替规律,通过林相改造,引入可自我更新青冈、红楠、苦槠和冬青等上海地区自然分布的物种,加速恢复进程,诱导群落进入进展演替阶段,从而实现人工林的自然恢复;而林下伴生物种如海桐、蚊母和小蜡等种源较丰富且可以自我更新,无需人为再次引入,同时要适当减少人工林的管护强度,制定相应的林相改造计划,从而建设群落结构完整、物种多样性丰富、生物量高、趋于稳定状态的少人工管护的“近自然型”绿地[31].
[1]王娟,达良俊,李艳艳,等.城市化进程中上海植被的多样性、空间格局和动态响应(Ⅴ):管护放弃后城市水杉林林下植被自然演替格局的研究[J].华东师范大学学报:自然科学版,2009(6):12-22.
[2]韩有志,王政权.森林更新与空间异质性[J].应用生态学报,2002,13(5):615-619.
[3]黄忠良,彭少麟,易俗.影响季风常绿阔叶林幼苗定居的主要因素[J].热带亚热带植物学报,2001,9(2):123-128.
[4]宋垚彬,张奇平,达良俊.浙江天童受损常绿阔叶林实验生态学研究(Ⅳ):土壤子库在不同受损常绿阔叶林恢复初期的作用[J].华东师范大学学报:自然科学版,2010(3):1-9.
[5]GUO C S,MING J Y,XIN H U,et al.Species-area relationship explained by the joint effects of dispersal limitation and habit heterogeneity[J].Ecology,2009,90(11):3033-3041.
[6]王传华,李俊清,陈芳青,等.鄂东南低丘地区枫香林下枫香幼苗更新限制因子[J].植物生态学报,2011,35(2):187-194.
[7]柴一新,祝宁.哈尔滨城市森林的天然更新[J].东北林业大学学报,2010,38(3):8-10.
[8]秦新生,刘苑秋,邢福武.低丘人工林林下植被物种多样性初步研究[J].热带亚热带植物学报,2003,11(3):223-228.
[9]康冰,王得祥,崔宏安,等.秦岭山地油松群落更新特征及影响因子[J].应用生态学报,2011,22(7):1659-1667.
[10]达良俊,杨永川.浙江天童国家森林公园常绿阔叶林主要组成种的种群结构及更新类型[J].植物生态学报,2004,28(3):376-384.
[11]熊小刚,熊高明,谢宗强.神龙架地区常绿落叶阔叶混交林树种更新研究[J].生态学报,2002,22(11):2001-2005.
[12]程积民,赵凌平,程杰.子午岭60年辽东栎种子质量与森林更新[J].北京林业大学学报,2009,31(2):10-16.
[13]金莹杉,翟明普,马润国,等.北京西山风景林林下更新种空间分布特征研究[J].西北林学院学报,2007,22(3):11-15.
[14]李小双,鹏明春,党承林.植物自然更新研究进展[J].生态学杂志,2007,26(12):2081-2088.
[15]达良俊,杨同辉,宋永昌.上海城市生态分区与城市森林布局研究[J].林业科学,2004,40(4):84-88.
[16]达良俊,杨永川,陈燕萍.上海大金山岛的自然植物群落多样性研究[J].城市林业,2004,2(3):22-25.
[17]方和俊.上海城市绿地植物群落现状及综合评价研究[D].上海:华东师范大学,2006.
[18]宋永昌.对中国植被分类系统的认知和建议[J].植物生态学报,2011,35(8):882-892.
[19]任海,王俊.试论人工林下乡土树种定居限制问题[J].应用生态学报,2007,18(8):1855-1860.
[20]GODEFROID S,RUCQUOLJ S,KOEDAM N.To what extent do forest herbs recover after clearcutting in beech forest[J].Forest Ecology and Management,2005,2(10):39-53.
[21]PAZ H,MAZER S J,MARTINEZ RAMOS M.Comparative ecology of seed mass in Paychotria(Rubiaceae):Within and between species effects of seed mass on early performance[J].Functional Ecology,2005,19(4):707-718.
[22]王娟.上海中心城区人工植被自然恢复潜力研究[D].上海:华东师范大学,2009.
[23]任海,彭少麟,陆宏芳.退化生态系统恢复与恢复生态学[J].生态学报,2004,24(8):1756-1764.
[24]HARPER J L.The Population Biology of Plants[M].New York:Academic Press,1977.
[25]彭少麟.南亚热带森林群落动态学[M].北京:科学出版社,1997.
[26]汤景明,翟明普.影响天然林树种更新因素的研究进展[J].福建林学院学报,2005,25(4):379-383.
[27]王瑞.上海市环城林带人工植被更新格局与恢复潜力研究[D].上海:华东师范大学,2010.
[28]臧润国,刘静艳,董大方.林隙动态与森林生物多样性[M].北京:中国林业出版社,1999.
[29]徐振邦,代力民,陈吉泉,等.长白山红松阔叶混交林森林天然更新条件的研究[J].生态学报,2001,21(9):1413-1420.
[30]高贤明,王巍,杜晓军,等.北京山区辽东栎林的径级结构、种群起源及生态学意义[J].植物生态学报,2001,25(6):673-678.
[31]达良俊,杨永川,陈鸣.生态型绿化法在上海“近自然”群落建设中的应用[J].中国园林,2004,3(38):38-40.
猜你喜欢
林业勘查设计(2022年1期)2022-02-15
乡村科技(2021年17期)2021-10-20
安顺学院学报(2021年4期)2021-09-16
林业调查规划(2020年3期)2020-06-03
现代园艺(2017年19期)2018-01-19
现代园艺(2017年23期)2018-01-18
现代园艺(2017年21期)2018-01-03
现代园艺(2017年21期)2018-01-03
新农业(2016年17期)2016-08-16
山东林业科技(2016年5期)2016-07-05
