
猴免疫缺陷病毒衣壳蛋白p27在大肠杆菌中的表达及纯化
2013-11-27 05:27马兰芝白杰英赵彦斌王洪宝丁璐静武文卿孙兆增
中国比较医学杂志 2013年1期
李 敏,马兰芝,白杰英,刘 一,赵彦斌,王洪宝,丁璐静,武文卿,孙兆增,曾 林
(1.军事医学科学院实验动物中心,北京 100071;2.军事医学科学院科技部,北京 100850)
猴免疫缺陷病毒(simian immunodeficiency virus,SIV)又名猴艾滋病病毒,是猴获得性免疫缺陷综合征(simianaquiredimmunedeficiency syndrome,SAIDS)的致病病毒[1]。研究人员于1985年第一次从饲养的猕猴体内分离到 SIV[2-4]。SIV感染主要破坏免疫系统,可导致细胞免疫和体液免疫缺陷。猕猴感染SIV可能会在其感染后期发展成为猴艾滋病(SAIDS),SAIDS是一种慢性传染病,致死率高,严重威胁猴群健康。因此,及时、准确的检测出SIV,并控制其传播,对于猴群的健康及生产具有重要意义。
SIV与人类免疫缺陷病毒(humanimmunodeficiency virus,HIV)基因同源性高,其形态、理化特性、分子生物学特性和致病机制均与HIV十分相似[5]。SIV感染猴可以作为研究人类艾滋病的理想动物模型,用于艾滋病的各项研究[6,7]。在模型的构建中,需检测 SIV 的病毒水平来进行评价。
衣壳蛋白 p27[8],是 SIV病毒颗粒的主要结构蛋白,其氨基酸序列高度保守,并含有诱导机体体液免疫和细胞免疫的主要抗原决定簇,被认为是SIV抗原检测的标志物[1]。因此,获得 p27纯化蛋白对SIV检测方法的建立具有重要意义[9]。国内已有利用 p27进行检测试剂盒的研究[10-12],但目前还未见商品化的p27抗原检测试剂盒的报道。
本研究利用基因合成方法,合成SIV衣壳蛋白p27的特异性抗原表位基因,成功构建了 pMAL-p5x-p27重组质粒,诱导其在大肠杆菌中高效表达,并对所表达蛋白进行纯化,以期为 SIV检测试剂盒的研制奠定基础。
1 材料和方法
1.1 材料
大肠杆菌DH5α、BL21(DE3),表达载体 pMAL-p5x由本实验室保存;限制性核酸内切酶、T4DNA连接酶为宝生物公(TaKaRa)产品;质粒提取试剂盒为QIAGENE公司产品;PCR产物回收试剂盒为QIAGENE公司产品;DNA分子量标准为康为世纪公司产品;蛋白质分子量标准为康为世纪公司产品;葡萄糖为国产分析纯试剂;IPTG为进口分析纯试剂,Amylose Resin亲和层析柱为NEB公司产品。
1.2 方法
1.2.1 基因合成
从GenBank上获得SIV的p27基因序列,利用生物信息学软件选择衣壳蛋白p27抗原表位集中的区域,并在基因序列5‘和3’端分别添加 NdeⅠ和EcoRⅠ酶切位点;该基因片段由上海捷瑞生物工程有限公司合成,合成序列全长753 bp,命名为p27。
1.2.2 重组表达质粒的构建
将合成的含有p27基因的质粒和表达载体质粒pMAL-p5x同时用 NdeⅠ和 Eco RⅠ进行双酶切,回收双酶切片段;用T4 DNA连接酶进行连接,构建重组质粒pMAL-p5x-p27;重组质粒转化BL21感受态细胞,提取质粒进行PCR及酶切鉴定,筛选阳性重组质粒。
1.2.3 p27重组蛋白的诱导表达
取阳性单菌接种于含有氨苄及10 mmol/L葡萄糖的LB培养液,37℃培养至至OD值为0.4~0.6,加入终浓度为0.3 mmol/L的 IPTG,30℃诱导4 h,离心收集菌体,进行10%SDS-PAGE分析。
1.2.4 p27重组蛋白的纯化
将1 L上述诱导后收集的表达菌体,用25 mL柱上样缓冲液重悬,-20℃过夜,第2天将其溶化后,置于冰浴中进行超声破碎20 min(15 min(超声5 s,间歇 15 s);4℃ 20000 r/min 离心 20 min,收集上清;采用 Amylose Resin亲和层析柱纯化目的蛋白,用包含10 mmol/L麦芽糖的上柱缓冲液进行洗脱,收集纯化蛋白,进行10%SDS-PAGE电泳分析。
2 结果
2.1 合成基因的鉴定
p27合成基因的PCR扩增产物经琼脂糖凝胶电泳显示,PCR产物大小约为750 bp(见图1),与p27基因相符,证实合成基因正确。
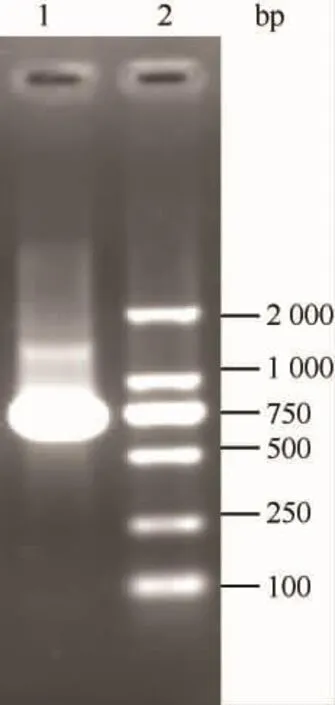
图1 p27 PCR产物琼脂糖凝胶电泳。Fig.1 Amplication of p27 DNA fragment by PCR.
2.2 重组表达质粒的鉴定
重组表达质粒用NdeⅠ和Eco RⅠ双酶切,经琼脂糖凝胶电泳显示,可见约为5700 bp和750 bp2条DNA条带(见图2),一条为 p27目的基因片段,约750 bp;另一条为pMAL-p5x载体基因片段,大小约为5700 bp。经测序鉴定,目的基因序列与预期相符。

图2 pMAL-p5x-p27重组质粒的酶切鉴定。Fig.2 Restriction map of recombinant plasmid.
2.3 p27重组蛋白的诱导表达
pMAL-p5x-p27融合蛋白经10%SDS-PAGE电泳分析,可见约70×103的明显条带(见图3),与预期重组蛋白分子质量大小一致。
2.4 p27重组蛋白的纯化
诱导产物经超声波破碎获得的上清用麦芽糖结合蛋白亲和层析柱纯化,经10%SDS-PAGE电泳分析,可见获得纯的p27重组蛋白(见图4)。
3 讨论
选择重组蛋白表达系统时,需要考虑表达产物的产量、可溶性、纯化效果等。本试验选用可以高效融合表达的pMAL-p5x原核表达载体,这种载体系统可以产生麦芽糖结合蛋白融合的重组蛋白,增强了重组蛋白的可溶性。并且本试验对诱导表达温度、时间和 IPTG浓度进行优化,选择用浓度为0.3 mmol/L IPTG在30℃低温诱导4 h来增加蛋白的表达量和可溶性。试验结果证明表达的p27融合蛋白表达量高,且多以可溶形式存在,为表达蛋白的纯化提供了便利。本试验采用麦芽糖结合蛋白亲和层析柱纯化方法,此法操作简单、经济方便,纯化效果好。经纯化获得目的蛋白纯度可达90%,纯化获得的p27融合蛋白为诊断试剂的研发奠定了基础。
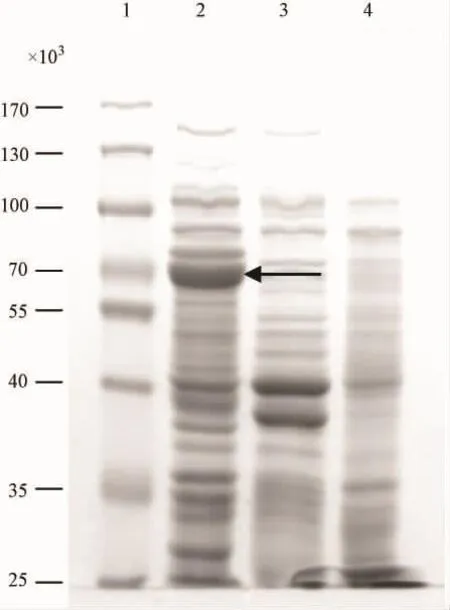
图3 表达产物的10%SDS-PAGE电泳。Fig.3 10%SDS-PAGE analysis of expressed products.
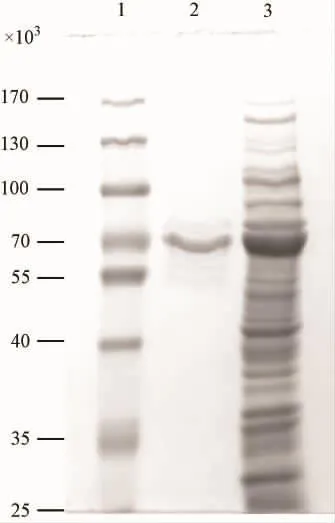
图4 融合蛋白纯化后的10%SDS-PAGE电泳。Fig.4 10%SDS - PAGE analysis of purified product.
SIV的衣壳蛋白 p27氨基酸序列比较保守[8]。利用p27抗原和p27抗体,可以在细胞和体液水平对SIV感染猕猴动物模型进行研究,对艾滋病的相关机制研究、疫苗研发及其免疫策略有效性进行评价[13]。因此,本试验中我们选取SIV的p27基因来进行研究。
本试验根据NCBI已发表SIV p27基因序列,设计合成p27基因;应用DNA重组技术,成功构建出重组质粒 pMAL-p5x-p27;完成了重组质粒 pMAL-p5x-p27在大肠杆菌BL21(DE3)中的高效表达。综上所述,本研究为SIV检测试剂盒的研究及应用奠定了一定基础。
[1] 殷震,刘景华,等.动物病毒学[M].第2版.北京:科学出版社,1997.
[2] Letvin N, Eaton K, Aldrich W, et al. Acquired immunodeficiency syndrome in a colony of macaque monkeys[J].Proceedings of the National Academy of Sciences of the United States of America.1983,80(9):2718-2722.
[3] Daniel MD,Letvin NL,King NW ,et al.Isolation of T-cell tropic HTLV-III-like retrovirus from macaques[J].Science.1985,228(4704):1201-1204.
[4] King NW,Hunt RD,Letvin NL.Histopathologic changes in macaques with an acquired immunodeficiency syndrome(AIDS)[J].The American journal of pathology.1983,113(3):382-388.
[5] KodamaT, Burns DP, Silva DP, etal. Strain-specific neutralizing determinant in the transmembrane protein of simian immunodeficiency virus[J].J Virol,l991,65(4):2010-2018.
[6] Miller CJ,Li Q,Abel K,et al.Propagation and dissemination of infection after vaginal transmission of simian immunodeficiency virus[J].J Virol,2005,79(14):9217 - 9227.
[7] Nehete PN,Nehete BP,Manuri P,et al.Protection by dendritic cells based HIV synthetic peptide cocktail vaccine:preclinical studies in the SHIV-rhesus model[J].Vaccine,2005,23(17- 18):2154- 2159.
[8] Amara RR,Patel K,Niediziela G,et al.A combination DNA and attenuated simian immunodeficiency virus vaccine strategy provides enhanced protection from simian/human immunodeficiency virus-induced disease[J].J Virol,2005,79(24):15356-15367.
[9] Triques K, Bourgeois A,Vidal N,et al. Near-full-length genome sequencing of divergent African HIV type I subtype F viruses leads to the identification of a new HIV type I subtype designated[J].AIDS Res Hum Retroviruses,2000,16:139-151.
[10] 季芳,刘晓明,饶军华.猴免疫缺陷病毒(SIV)双抗原夹心ELISA诊断试剂盒的研制及应用[J].实验动物科学,2008,25(3):26-29.
[11] 孙敏,魏强,梁成珠,等.SIVp27抗原双抗体夹心 ELISA检测方法的研究[J].中国动物疫,2007,24(4):32 -35.
[12] 华秋东,姜成刚,王海,等.人/猴免疫缺陷病毒 SHIV衣壳蛋白p27的表达纯化及活性鉴定[J].东北农业大学学报,2008,39(11):51 -55.
[13] Buch S,Pinson D,Hou Y,et al.Neuropathogenesis of chimeric simian human immunodeficiency virus infection in rhesus macaques[J].J Med Primatol.2000,29(3 -4):96 -106.
猜你喜欢
成都医学院学报(2022年4期)2022-08-19
解放军医学院学报(2022年1期)2022-04-02
科学导报(2021年82期)2021-12-17
江西农业学报(2021年4期)2021-04-20
昆明医科大学学报(2021年1期)2021-02-07
三农资讯半月报(2020年11期)2020-06-21
医药前沿(2018年27期)2018-01-16
肿瘤影像学(2015年3期)2015-12-09
中国当代医药(2015年9期)2015-03-01
癌变·畸变·突变(2015年3期)2015-02-27
