
造纸废水对芦苇实生苗保护酶及叶绿素荧光的影响
2013-11-05 06:39孙景宽陆兆华
江西农业大学学报 2013年3期
孙景宽,陆兆华* ,李 田
(1.中国矿业大学(北京)恢复生态学研究所,北京100083;2.滨州学院 山东省黄河三角洲生态环境重点实验室,山东滨州256603)
近年来,由于自然因素和人为因素的影响,黄河三角洲湿地退化极其严重,充分利用一切可利用的水资源是恢复黄河三角洲退化湿地的关键。黄河三角洲一些造纸行业废水排放量大,有机物含量高,COD 浓度大,废水必须经处理达标后排放,在一定程度上增加了企业的生产成本,降低了企业的市场竞争能力[1]。
利用造纸废水修复退化芦苇湿地,既可以解决水资源缺乏的问题,又能解决造纸废水的排放问题,变废为宝,使造纸废水得到资源化利用。芦苇(Phragmites communis Trim.)为禾本科芦苇属多年生植物,是具有较强污水净化能力的湿地植物之一,因而常用于人工湿地污水处理系统中[2-6]。目前,关于废水对芦苇植株生长、生理、生态等效应的研究较多[7-11],但这些研究对象主要是根状茎盆栽芦苇和野外形成种群的芦苇。通过种子萌发形成实生苗是芦苇种群扩展繁殖的一个重要方式,利用造纸废水修复退化芦苇湿地,需要考虑造纸废水对芦苇实生苗的影响,但关于这方面的研究未见报道,本研究选用芦苇实生苗,进行不同浓度造纸废水的胁迫试验,揭示芦苇实生苗芦苇保护酶和叶绿素荧光对造纸废水的响应机制,探索芦苇实生苗对造纸废水的耐受能力,为造纸废水灌溉修复自然环境中的芦苇实生苗种群提供理论依据。
1 材料与方法
1.1 实验材料
芦苇(Phragmites communis Trim.)种子于种子成熟季节采自黄河三角洲自然生态系统,次年3 月于山东省黄河三角洲生态环境重点实验室数控温室育苗。
1.2 胁迫方法
播种30 d 后,将芦苇从土中挖出,洗净泥土,在水桶(盆直径为30 cm,高为25 cm)中水培,每桶4株,缓苗7 d 后,用不同浓度造纸废水开始胁迫处理,每3 d 更换1 次溶液。造纸废水来自山东海韵造纸厂,废水主要水质指标:化学需氧量(CODcr)900 mg/L,生化需氧量(BOD)84 mg/L,pH 7. 44,总氮28.4 mg/L,总磷7. 2 mg/L,矿化度2 080 mg/L,用自来水稀释成5 种不同浓度:CODcr浓度分别为75 mg/L,150 mg/L,300 mg/L,600 mg/L,900 mg/L,以自来水为对照(CK)。
1.3 测定方法
在胁迫30 d 后选取芦苇上部健康完全展开的叶片(顶端往下3 ~5 片),将其剪碎,从中称取一定量的叶片对相关指标进行测定,每一指标的测定3 次重复。
保护酶的提取:取0.3 g 叶片切段,置于预冷的研钵中,加适量的预冷的50 mmol/L 磷酸缓冲液(含1% 聚乙烯吡咯烷酮,pH 7.0)及少量石英砂,在冰浴中研磨成匀浆,将匀浆液全部转入到15 mL 离心管中,于2 ~4 ℃,12 000 g 离心20 min,上清液转入25 mL 容量瓶中,沉淀用5 mL 磷酸缓冲液再提取2次,上清液并入容量瓶中,定容到刻度,4 ℃下保存备用[12]。SOD 的测定以抑制氯化硝基四氮唑蓝光化还原50%为一个酶活性单位表示[13]。POD 的测定用愈创木酚染色法,以每分钟内A470 变化0.01 为一个过氧化物酶活性单位[13]。CAT 的测定用紫外吸收法,以每分钟内A240 减少0.1 的酶量为一个酶活性单位[14]。
MDA 的提取:取叶片0.2 g,加入10% TCA 2.0 mL 和少量石英砂,研磨;转移到离心管中,控制在10 mL 以内,4 000 ×g 离心10 min,定容到10 mL。即为样品提取液。MDA 测定和计算按照张志良等[13]的方法。
叶绿素荧光参数测定:在胁迫30 d 后,用OS-30P 叶绿素荧光仪测定。芦苇幼苗经过12 h 叶夹暗适应后,选取上部健康完全展开的叶片(顶端往下3 ~5 片)测定初始荧光(Fo),最大荧光(Fm),然后按公式Fv=Fm-Fo 计算出可变荧光(Fv),PSⅡ最大光化学量子产量(Fv/Fm)。
1.4 数据处理
用SPSS13.0 进行单因素方差分析和Duncan 多重比较。
2 结果与分析
2.1 芦苇叶片SOD、POD、CAT 活性对造纸废水的响应
SOD、POD、CAT 是植物体内清除自由基的重要保护酶[15]。SOD 在植物抵抗外界环境胁迫中起着非常重要的作用。SOD 能将超氧阴离子自由基清除氧化成H2O2和O2,POD、CAT 能将H2O2转变为H2O 和O2[16]。图1 显示了芦苇SOD 活性对不同浓度造纸废水的响应,从图中可以看出,随着造纸废水CODcr的增加,SOD 活性逐渐升高,在造纸废水CODcr为300 mg/L 达到最大值,为对照的2.81 倍,然后SOD 活性又逐渐下降。造纸废水CODcr为900 mg/L,SOD 活性为对照1.8 倍。这说明300 mg/L 为SOD 的耐受阈值,超过这个阈值,SOD 活性下降,但活性仍高于CK。图2 显示了芦苇POD 活性对不同浓度造纸废水的响应,从图中可以看出,芦苇在遭受废水胁迫后,其POD 活性迅速增加,在CODcr为600 mg/L 时POD 活性达到最高值,为对照的2.97 倍,在CODcr为900 mg/L 时,芦苇的POD 活性虽有所降低,但仍高于对照和75 mg/L。图3 显示了芦苇CAT 活性对不同浓度造纸废水的响应,从图中可以看出,当废水CODcr浓度为75 mg/L 时,芦苇CAT 活性和对照差异不显著,然后,随着废水浓度的增高,芦苇幼苗叶片中CAT 活性逐渐增加,废水CODcr浓度为900 mg/L 最大,此时CAT 活性为对照的2.8 倍,说明CAT 对造纸废水胁迫变化比较敏感。
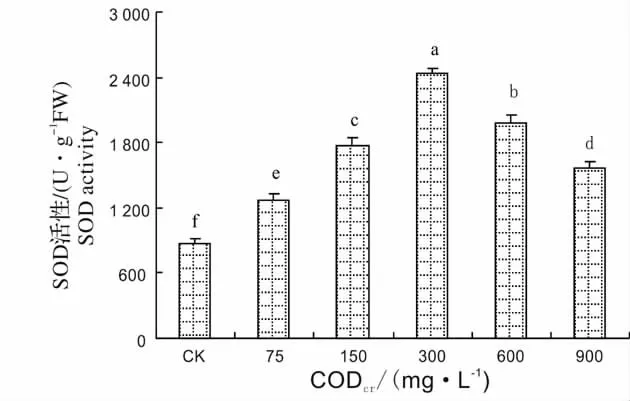
图1 芦苇SOD 活性对造纸废水的响应Fig.1 Response of SOD activity of Phragmites communis Trim.to papermaking wastewater
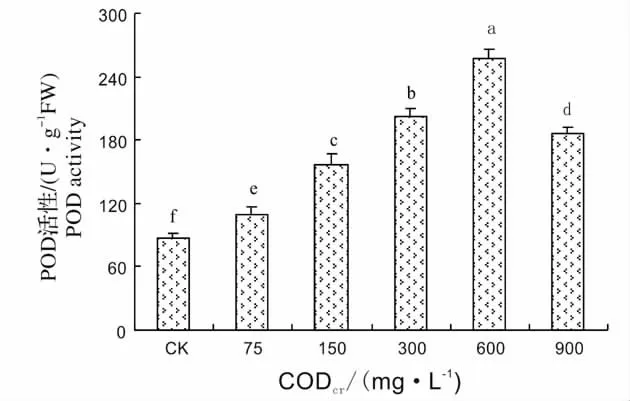
图2 芦苇POD 活性对造纸废水的响应Fig.2 Response of POD activity of Phragmites communis Trim.to papermaking wastewater
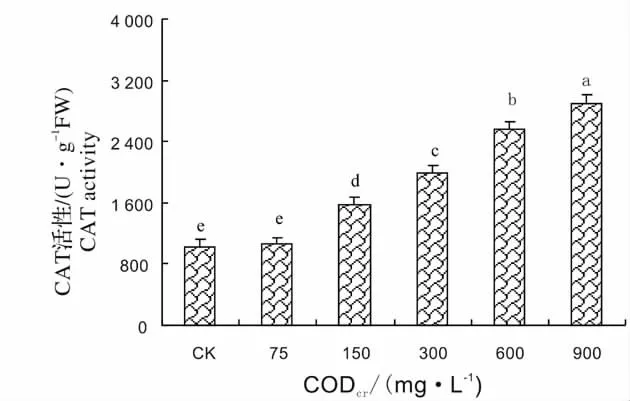
图3 芦苇CAT 活性对造纸废水的响应Fig.3 Response of CAT activity of Phragmites communis Trim.to papermaking wastewater
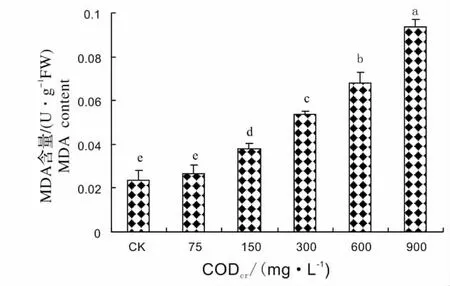
图4 芦苇MDA 含量对造纸废水的响应Fig.4 Response of MDA content of Phragmites communis Trim.to papermaking wastewater stress
2.2 芦苇叶片MDA 含量对不同浓度造纸废水的响应
丙二醛(MDA)是膜脂过氧化的重要产物,通常利用它作为膜脂过氧化指标,其含量的变化可反映逆境条件下膜系统受伤害的程度[7]。图4 显示了芦苇MDA 含量对不同浓度造纸废水的响应,从图中可以看出,当废水CODcr浓度为75 mg/L 时,芦苇MDA 含量和对照差异不显著,然后,随着废水浓度的增高,芦苇幼苗叶片中MDA 含量逐渐增加,废水CODcr浓度为900 mg/L 最大,此时MDA 含量为对照的3.98 倍。说明造纸废水浓度越高,芦苇膜质过氧化程度越严重。
2.3 造纸废水对芦苇叶绿素荧光的影响
初始荧光是光系统Ⅱ反应中心处于完全开放时的荧光产量。初始荧光的减少表明光合色素的热耗散增加,而增加则表明光系统Ⅱ反应中心受到一定程度的破坏[17]。在废水CODcr浓度为75 ~300 mg/L时,初始荧光和对照的差异不显著,CODcr浓度600、900 mg/L,初始荧光和对照的差异显著,说明此时芦苇光系统Ⅱ反应中心受到一定程度的破坏。最大荧光是光系统Ⅱ反应中心完全关闭时的荧光产量,反映了光系统Ⅱ的电子传递情况[18]。随着废水CODcr浓度的升高,芦苇叶片最大荧光呈现先增加后降低的趋势,在废水浓度为300 mg/L 时最高。光系统Ⅱ最大光化学效率反映光系统Ⅱ反应中心原初光能转化效率[19]。随着废水CODcr浓度的升高,芦苇叶片光系统Ⅱ最大光化学效率呈现先增加后降低的趋势,废水CODcr浓度为75 mg/L 时,光系统Ⅱ最大光化学效率和CK 的差异不显著,150 mg/L、300 mg/L时,光系统Ⅱ最大光化学效率显著高于CK,废水CODcr浓度为600 mg/L、900 mg/L 时,光系统Ⅱ最大光化学效率值显著低于对照,表明此时芦苇受到了光抑制。
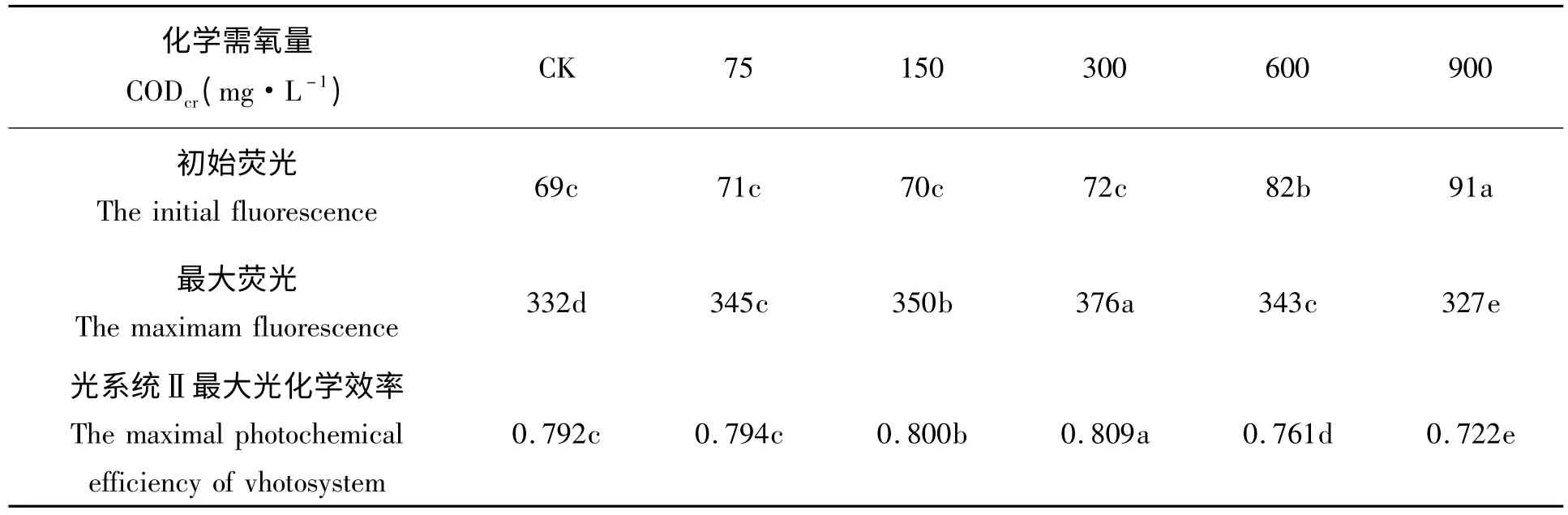
表1 造纸废水对芦苇叶绿素荧光特性的影响Tab.1 Response of chlorophyll fluorescence characteristics of Phragmites communis Trim.to papermaking wastewater stress
3 讨论
植物体内酶系统总的变化趋势是处于动态平衡的,当植物受到逆境胁迫时,在逆境条件下,植物体内会产生大量的自由基,SOD、POD、CAT 等保护酶在清除自由基的过程中发挥了重要作用[20]。肖德林等[8]对在工湿地生长的成年芦苇植株研究表明,两种浓度废水胁迫下,SOD 活性均明显上升,POD 活性在胁迫初始时急速下降,到中后期则逐步恢复到胁迫前的水平,CAT 活性则在整个处理期间逐渐下降,推测废水胁迫未能导致芦苇抗氧化酶系统启动,其保护机制可能是一些非酶促活性氧清除系统。本研究表明,随着造纸废水CODcr的增加,SOD、POD 酶活性呈现先升高后下降的趋势,
CAT 随着造纸废水CODcr的增加呈现持续升高趋势,这可能和它们是协同保护酶有关[21]。根据本研究结果,可以推测造纸废水胁迫导致了芦苇实生苗保护酶系统的启动,这和肖德林等[8]研究结果存在差异,究其原因还有待进一步研究。
1975 年,Fridovich 提出生物自由基伤害学说,认为植物体内自由基大量产生会引发膜脂过氧化作用,造成细胞膜系统破坏,严重时导致植物死亡,通常用膜质过氧化产物丙二醛的变化来衡量膜的完整性和膜质过氧化程度[22]。尽管保护酶在清除自由基的过程中发挥了重要作用,芦苇实生苗MDA 含量随着造纸废水CODcr的增加而逐渐增加,并且增加幅度随着造纸废水CODcr的增加而增大。这种变化趋势可能与SOD、POD 变化趋势相关,CAT 活性抑制MDA 的效果不显著。
叶绿素荧光与光合作用中各个反应过程紧密相关,任何逆境对光合作用各过程产生的影响都可通过体内叶绿素荧光诱导动力学变化反映出来,与一些“表观性”的气体交换指标相比,叶绿素荧光参数更反映“内在性”的特点,而且它具有快速、灵敏、非破坏性测量等优点,被视为研究植物光合作用与环境关系的内在探针[23]。本研究表明,废水CODcr浓度高于为300 mg/L时,初始荧光显著高于对照,最大荧光、光系统Ⅱ最大光化学效率显著低于对照,说明此时芦苇光系统Ⅱ反应中心受到一定程度的破坏,并且芦苇实生苗出现显著的光抑制。
综上所述,芦笋实生苗保护酶和叶绿素荧光参数对造纸废水具有一定的适应性,CODcr浓度为300 mg/L时,芦苇SOD 酶活性、最大荧光、光系统Ⅱ最大光化学效率最高,超过此废水浓度,芦苇叶片膜质过氧化程度加重,光系统Ⅱ反应中心受到一定程度的破坏。本实验只是研究了保护酶和叶绿素荧光参数对造纸废水的响应,今后应结合芦苇实生苗生长、光合、离子运移等,综合分析适宜浇灌芦苇实生苗的造纸废水CODcr浓度。同时深入开展田间实验,可为利用造纸废水恢复芦苇野外实生苗种群提供更重要的参考依据。
[1]李在田.芦苇人工湿地造纸废水资源化利用[J].水处理技术,2006,32(2):76 -77.
[2]苏芳莉,周欣,陈佳琦,等.芦苇湿地生态系统对造纸废水中铅的净化研究[J].中国环境科学,2011,31(5):168-773.
[3]刘倩,胡冲,谢冰,等.芦苇人工湿地对垃圾渗滤液总氮去除能力的研究[J].水处理技术,2010,36(12):39-46.
[4]Martin M K,Robert J F,Charles P G,et a1.Management of dairy waste in the sonoran desert using constructed wetland technology[J].Water Science and Technology ,1999,40(3):57- 65.
[5]吴湘,王友慧,郭建林,等.3 类水生植物对池塘养殖废水氮磷去除效果的研究[J].西北植物学报,2010,30(9):1876-1881.
[6]杨林,伍斌,赖发英,等.7 种典型挺水植物净化生活污水中氮磷的研究[J].江西农业大学学报,2011,33(3):616-621.
[7]邓仕槐,肖德林,李宏娟,等. 畜禽废水胁迫对芦苇生理特性的影响[J]. 农业环境科学学报,2007,26(4):1370 -1374.
[8]肖德林,邓仕槐,李宏娟,等.畜禽废水胁迫对芦苇叶绿素含量及抗氧化酶系统的影响[J].农业环境科学学报,2007,26(4):1370 -1374.
[9]孙翠玲,袁东.造纸尾水对芦苇生长和生态环境的影响[J].中国环境管理干部学院学报,2005,15(2):82 -85.
[10]丁成,王世和,杨春生.草浆废水灌溉对海涂湿地土壤及芦苇生长的影响[J].生态环境,2005,14(1):21 -25.
[11]任珺,陶玲,罗青龙,等.污水环境对芦苇生长性状的影响[J].环境科学与技术,2010,10(1):158 -161.
[12]李柏林,梅慧生.燕麦叶片衰老与活性氧代谢的关系[J].植物生理学报,1989,15(1):6 -12.
[13]张志良,瞿伟菁.植物生理学实验指导[M].北京:高等教育出版社,2003.
[14]Trevor E,Kraus R,Austin F.Paclobutrazol protects wheat seedlings from heat and paraquat injury is detoxification of active oxygen involved[J].Plant Cell Physiol,1994,35:45 -52.
[15]Noctor G,Foyer C H.Ascorbate and glutathione:Keeping active oxygen under control[J].Annual Review of Plant Physiology and Plant Molecular Biology,1998,49(1):249 -279.
[16]Chaitanya K V,Sundar D,Masilamani S,et al.Variation in heat stress -induced antioxidant enzyme activities among three mulberry cultivars[J].Plant Growth Regulation,2002,36(2):175 -180.
[17]Demmig B,Bjorkman O.Comparison of the effect of excessive light on chlorophyll fluorescene (77K)and photon yield of O2evolution of leaves of higher plants[J].Planta,1987,17l:l71 -l84.
[18]Ji X B,Holloeher T C.Reduction of nitrite to nitric oxide by enteric bacteria[J].Biochem Biophys Res Conunun,1988,157:106 -108.
[19]张守仁.叶绿素荧光动力学参数的意义及讨论[J].植物学通报,1999,16(4):444 -448.
[20]Gina Brito,Armando Costa,Henrique M A C,et al.Response of Olea europaea ssp.Maderensis in vitro shoots exposed to osmotic stress[J].Scientia Horticu Nurae,2003,97:411 -417.
[21]孙景宽,夏江宝,田家怡,等.干旱胁迫对沙枣幼苗根茎叶保护酶系统的影响[J].江西农业大学学报,2009,31(5):1011 -1016.
[22]Sundar D,Perianayaguy B,Reddy A R.Localization of antioxidant enzymes in the cellular compartments of sorghum leaves[J].Plant Growth Regulation,2004,44(2):157 -163.
[23]崔晓伟,高健,张志坚,等.5 种地被竹叶绿素荧光特性研究[J].江西农业大学学报,2011,33(4):726 -730.
猜你喜欢
海外文摘·文学版(2022年4期)2022-04-14
农业科技通讯(2022年1期)2022-01-17
黄河之声(2021年19期)2021-02-24
阅读(科学探秘)(2020年8期)2020-11-06
山东林业科技(2019年2期)2019-06-03
中国果业信息(2019年1期)2019-01-05
——以新源县为例
新疆农业科技(2018年5期)2018-05-07
岁月(2018年2期)2018-02-28
中国诗歌(2017年12期)2017-11-15
生物学教学(2017年9期)2017-08-20
