
苏云金芽胞杆菌cry7Ab9基因的克隆与杀虫活性的测定
2013-10-11 11:22李海涛曲步云赵世源贾宏博高继国
湖南师范大学自然科学学报 2013年2期
李海涛,曲步云,赵世源,贾宏博,高继国
(东北农业大学生命科学学院,中国哈尔滨 150030)
苏云金芽胞杆菌(Bacillus thuringiensis,简称Bt)是革兰氏阳性菌,芽胞杆菌属的一个种[1].它形成芽胞的过程中,形成一个或多个伴胞晶体(parasporal crystal).大多数这种伴胞晶体具有杀虫活性,所以又称为杀虫晶体蛋白(ICPs)或δ-内毒素(δ-endotoxin).Bt蛋白杀虫活性对非靶标生物无害,因此被制成生物杀虫剂广泛地应用于农业、林业以及卫生害虫的防治[2-3].
cry7类基因编码蛋白相对分子质量大约为130 000,形成菱形晶体[4].cry7类基因对鳞翅目、鞘翅目和直翅目害虫的生物防治方面有着广阔的前景,目前只报道了5种cry7类模式基因,cry7Aa、cry7Ab、cry7Ba、cry7Ca 和 cry7Da(http://www.lifesci.sussex.ac.uk/home/Neil_Crickmore/Bt.).其中 cry7Aa 杀马铃薯甲虫(LC50 为13.1 mg·L-1)[5],cry7Ba1 杀小菜蛾(3 天 LC50 为95.7 μg·L-1,5 天 LC50 为39.9 μg·L-1)[6],cry7Ca1 杀蝗虫(48 h LC50 为4.73 mg·L-1,72 h LC50 为 3.19 mg·L-1)[7].目前已经发现了 8 个 cry7Ab类基因,但只有邓淑等报道了cry7Ab4对鞘翅目叶甲科的大猿叶甲有一定的杀虫活性,而对甜菜夜蛾、小菜蛾无明显杀虫活性[8].鉴于cry7类基因有一定的研究价值,因此发掘新型cry7类基因,深入研究cry7类基因具有一定的实际意义[9].
1 材料与方法
1.1 材料
1.1.1 菌株与质粒 本文所用菌株和质粒见表1.

表1 菌株与质粒Tab.1 The strains and the plasmids
1.1.2 培养基 液体LB,固体LB,液体1/2LB,固体1/2LB.
1.1.3 试剂 限制性内切酶及连接酶、pfu购自GiBcol、TaKaRa公司.dNTP、Taq酶等购自生工生物工程公司.分析纯化学试剂均为市售.
1.1.4 试虫 暗黑鳃金龟幼虫(一龄,8~10 d),由中国农科院植保所和沧州植物保护研究所提供.
1.2 方法
1.2.1 cry7基因的鉴定与克隆 采用cry7类基因型的通用引物对本实验室分离得到的22株野生菌进行了PCR鉴定,通过K5un7/K3un7筛选挑取阳性克隆进行酶切分析和核苷酸序列测定[10].获取cry7类基因,将扩增目的片段连接PMD18-T载体,并进行测序.
1.2.2 cry7Ab全长基因的克隆 利用Bt已公布的cry7Ab类基因的全长序列,设计出用于扩增全长序列的引物,其中在5'端引物加进Bam H1酶切点,3'端引物加进SalⅠ酶切点,序列如下:
Q5UN7:5'-CGCGGATCCATGAATTTAAATAATTTAGGT-3'
Q3UN7:5'-ACGCGTCGACACATAGCTCTTCCATCAAAA-3'
用引物扩增得到cry7Ab 3.4 kb的全长基因,利用抗性筛选、PCR鉴定及酶切分析,筛选出含cry7Ab基因的阳性转化子[10-11].
1.2.3 cry7Ab9基因的原核表达及杀虫蛋白质的提取 将鉴定得到的重组质粒pEB-cry7Ab转化到大肠杆菌BL21(DE3)中,并进行阳性菌株筛选,同时利用IPTG诱导表达,收集诱导物离心弃上清,进行SDS-PAGE电泳进行检测.将获得的阳性菌株,诱导表达.利用10 mmol/L Tris-Cl(pH8.0)悬浮,超声波破碎菌体,收集上清,确定蛋白质浓度.
1.2.4 杀虫活性测定
将待测样品稀释成不同梯度,取适量干土豆丝加入待测样品稀释液浸泡再称取100 g试验用土混匀,分装入经消毒的6孔培养板中.然后每空接入暗黑鳃金龟幼虫一头,每个浓度梯度重复5次,放置25℃光照培养箱中,分别于7 d及两周调查死、活虫数.以10 mmol/L Tris-Cl(pH8.0)溶液作为对照,试验方法同上.
2 结果与分析
2.1 Bt QZG121菌株伴孢晶体形态的观察
Bt菌株QZG121在1/2LB培养基上培养48 h后,形成单菌落,菌落乳白色,圆形或近圆形,36 h左右可观察到晶体.挑取单菌落少量,制片,油镜下观察,可见伴胞晶体形态为菱形,见图1.
2.2 cry7基因的鉴定与克隆
对本实验室保存的22株野生菌利用K5un7/K3un7进行了cry7基因鉴定,发现一株含有1.3 kb左右片段的菌株QZG121.利用PMD18-T载体,筛选挑取阳性克隆,并对阳性克隆PMD18-T载体进行酶切分析及核苷酸序列测定.测序结果利用NCBI blast比对分析,确定核苷酸序列与cry7Ab最为相似,相似度最高达到96%.酶切鉴定结果见图2.
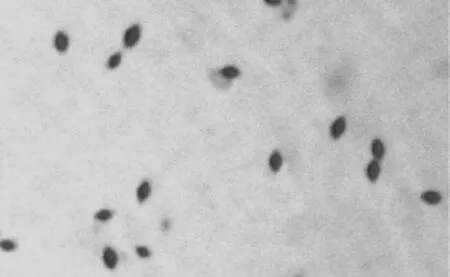
图1 Bt QZG121菌株的晶体形态特点Fig.1 The crystal form of Bt QZG121
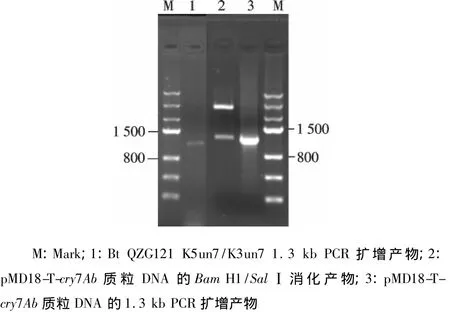
图2 基因的酶切与PCR鉴定分析Fig.2 Cry7Ab gene restriction and PCR analysis
2.3 cry7Ab全长基因的克隆
2.3.1 pEB-cry7Ab载体的构建 利用全长引物,进行全长基因的克隆表达,阳性转化子酶切分析及PCR鉴定结果,可以得到大小为5.7 kb的载体片段和3.4 kb cry7Ab基因片段,该cry7Ab全长基因表达载体构建正确.
2.3.2 cry7Ab全长序列及同源性分析 将获得的重组质粒pEB-cry7Ab测序,由DNA序列推导的氨基酸序列结果.cry7Ab蛋白由1 138个氨基酸组成,相对分子质量为129 437.其中亮氨酸Leu(L)、天冬酰胺Asn(N)、丝氨酸Ser(S)、异亮氨酸Ile(I)、谷氨酸Glu(E)较多,分别为 8.79%、7.38%、7.38%、7.12%、7.02%.该蛋白等电点 pH 5.045,为弱酸性蛋白[12].
该基因在 GenBank登记,其 Accession number为GU936713.并由Bacillus thuringiensis命名委员会命名为cry7Ab9,NCBI Blast比对该基因蛋白序列与cry7Ab6最相似(见表2),同源性为98.25%.
通过NCBI Conserved Domain分析结果表明该蛋白的DomainⅠ在氨基端第59~280位,由221氨基酸组成;DomainⅡ在第286~487位,合计201个氨基酸;DomainⅢ在第497~637位,共140个氨基酸组成.因此,Cry7Ab9蛋白质的生物杀虫活性区估计在氨基端59~637.用ANRHWETORV5.0软件对cry7Ab9进行蛋白质二级结构预测[13-14],结果表明,在其蛋白质二级结构中,40%为 α-螺旋,29%为 β-片层,无 β-转角,31%为无规则线圈.
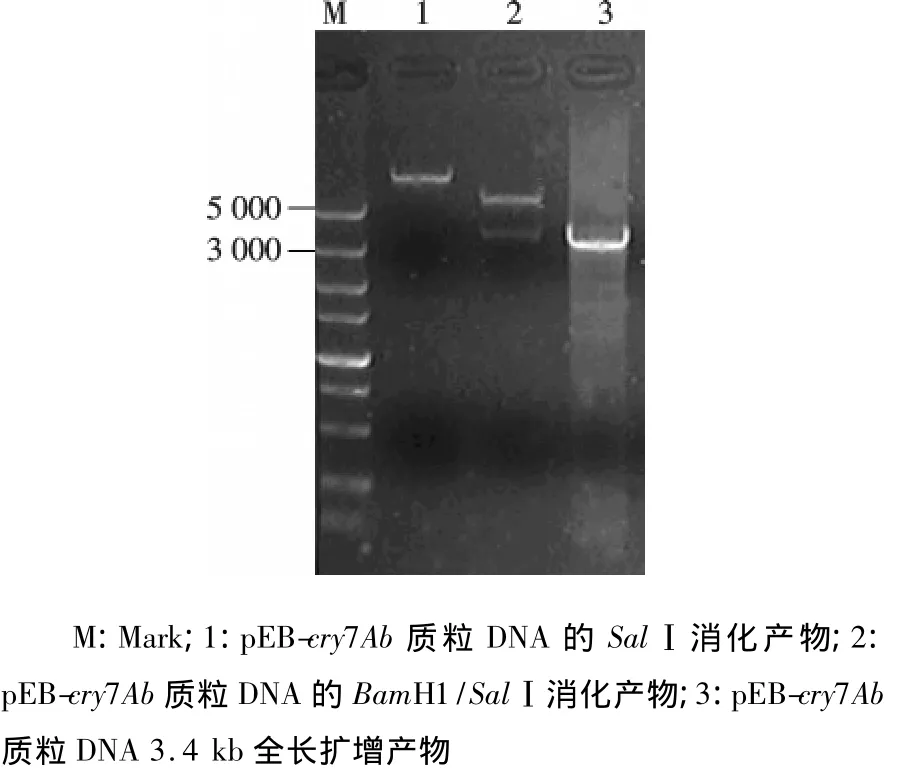
图3 pEB-cry7Ab的酶切与PCR鉴定分析Fig.3 pEB-cry7Ab restriction and PCR detection analysis
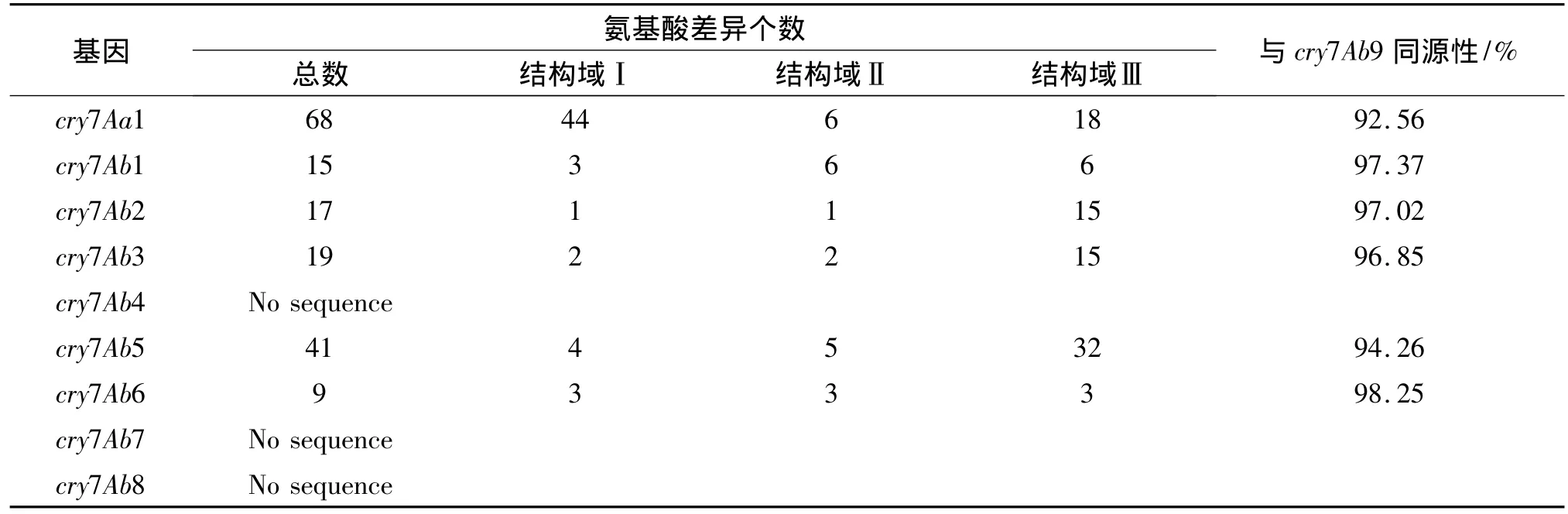
表2 Cry7Ab9与其他cry7Ab氨基酸的差异Tab.2 Difference of amino acid sequence between cry7Ab9 and other cry7Ab
2.4 Cry7Ab9 基因在 E.coli BL21(DE3)中的表达
利用pEB-cry7Ab9阳性质粒转化到大肠杆菌BL21(DE3)中,IPTG诱导表达.如图4所示电泳条带2在相对分子质量为130 000处有高效表达.结果表明,cry7Ab9基因能在大肠杆菌中高效表达相对分子质量为130 000的蛋白.
2.5 暗黑鳃金龟幼虫生测结果及数据分析处理
用大肠杆菌表达蛋白pEB-7Ab9进行蛴螬生物测定.生测为每个梯度3个重复,每个重复10头虫共30头虫,如表3所示.7天校正死亡率 10 mg·L-1为11.11%、100 mg·L-1为 25.93%;两周校正死亡率 10 mg·L-1为 33.33%、100 mg·L-1为 48.15%.计算LC50 为289.99 mg·L-1.结果表明,cry7Ab9 蛋白对暗黑鳃金龟幼虫显示了一定的杀虫活性.
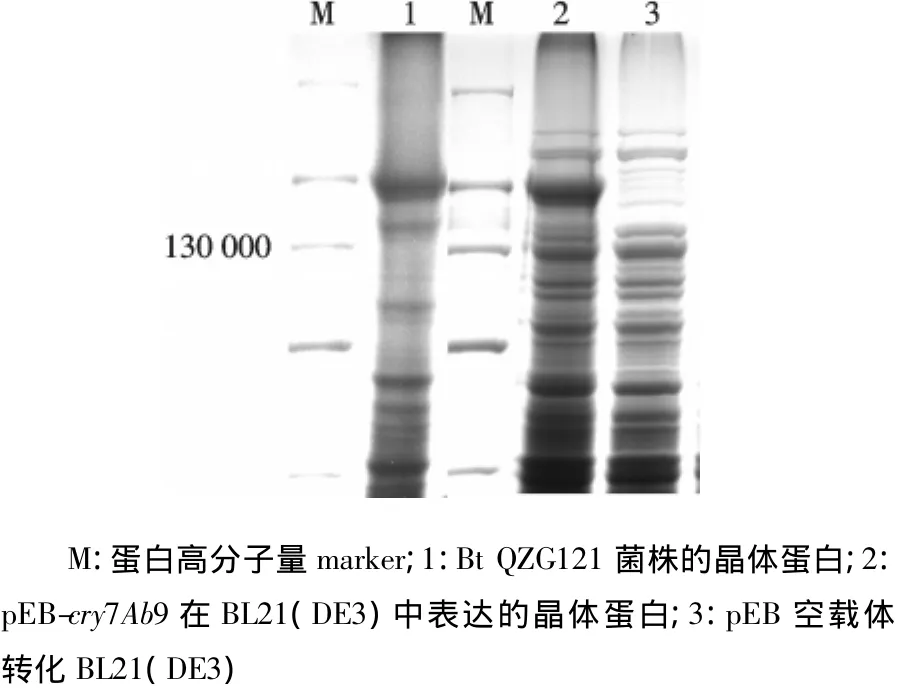
图4 Cry7Ab9在BL21(DE3)中的表达Fig.4 The cry7Ab9 protein expressed in BL21(DE3)
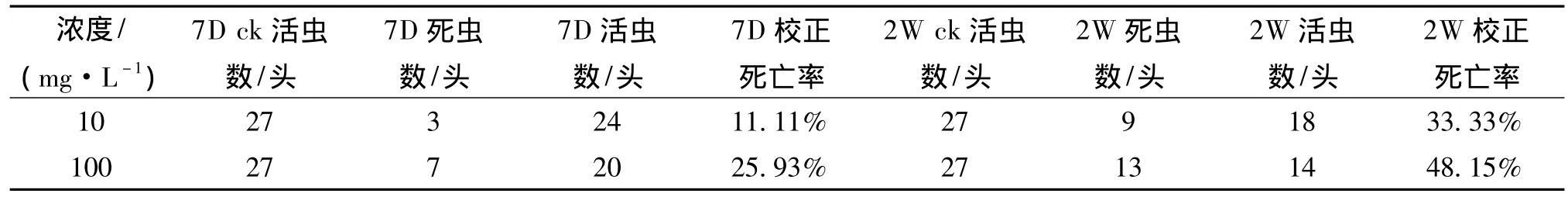
表3 暗黑鳃金龟幼虫生测结果Tab.3 The result of bioassay for Holotrichia parallela
3 讨论
本研究从千山分离得到22株苏云金芽胞杆菌中鉴定得到2株含有cry7基因型的菌株,检出率为9.09%.应用PCR法鉴定出其中一株Bt菌株QZG121中含有cry7Ab基因,并成功克隆到1个新的苏云金芽胞杆菌杀虫晶体蛋白基因cry7Ab9.通过构建原核表达载体,获得阳性重组质粒pEB-cry7Ab,转入E.coli BL21(DE3),cry7Ab9基因在E.coli BL21(DE3)中能正常表达.SDS-PAGE分析表明,表达蛋白的相对分子质量为130 000.目前只报道了5种cry7类模式基因,其中cry7Aa对马铃薯甲虫有杀虫活性,cry7Ba1对小菜蛾有杀虫活性,cry7Ca1对蝗虫有杀虫活性.它对鳞翅目、鞘翅目和直翅目害虫均具有较高杀虫活性,故cry7类基因存在巨大的挖掘潜力,在生物防治方面有着广阔的前景.本研究发现的cry7Ab9基因与已知的cry7Ab6基因相似性高达98.25%.目前已知的8种cry7Ab类基因,只有邓淑等报道cry7Ab4表达蛋白质对鞘翅目害虫有杀虫活性[8].本研究发现的cry7Ab9基因表达蛋白对暗黑鳃金龟幼虫进行初步生测,表现出一定杀虫活性.cry7Ab9新基因的发现不仅丰富了cry7类基因的种类,而且可以为鳞翅目、鞘翅目和双翅目害虫的防治以及抗性治理提供新的途径,也为研究蛋白质结构和功能的关系提供实验材料,特别是对鞘翅目类害虫的生物防治提供了新的基因资源.cry7Ab9基因与已知的cry7Ab6基因的相似性极高,但cry7Ab6基因并未表现出对鞘翅目的杀虫活性,这种相似性极高的同类基因所表现的活性差异,可以从生物信息学角度探索Bt杀虫晶体蛋白杀虫谱窄的解决方法,以期拓宽杀虫谱,提高Bt杀虫晶体蛋白在生物防治方面的利用价值.
[1]布坎南 R E,吉布斯 N E.伯杰氏细菌鉴定手册[M].北京:科学出版社,1984.
[2]SCHNEPF E,CRICKMORE N,Van RIE J,et al.Bacillus thuringiensis and its pesticidal crystal proteins[J].Microbiol Mol Biol R,1998,62(3):775-806.
[3]BRAVO A,GILL S S,SOBERÓN M.Mode of action of Bacillus thuringiensis Cry and Cyt toxins and their potential for insect control[J].Toxicon,2007,49(4):423-435.
[4]宋 萍,郭丽伟,苏俊平,等.含cry7类基因的苏云金芽胞杆菌菌株的分析[J].华北农学报,2010,25(4):40-43.
[5]BART L,HERMAN H,KATRIEN A,等.Novel Bacillus thuringiensis insecticidal crystal protein with a silent activity against coleopteran larvae[J].Appl Environ Microbiol,1992,58(3):2536-2542.
[6]张振宇.苏云金芽胞杆菌新异杀虫晶体蛋白基因的克隆与基因工程菌WG-001的安全评估[D].武汉:华中农业大学,2005.
[7]SONG L L,GAO M Y,DAI S Y,et al.Specific activity of a Bacillus thuringiensis strain against Locusta migratoria manilensis invertebrate pathology[J].J Inver Pat,2008,98(2):169-176.
[8]邓 淑,束长龙,林 毅,等.新型cry7Ab基因的鉴定、克隆、表达与杀虫活性[J].农业生物技术学报,2009,17(5):908-913.
[9]王立光.苏云金杆菌cry7Ab7基因的克隆表达及融合基因和工程菌构建[D].保定:河北农业大学,2010.
[10]张 杰,宋福平.PCR技术与ICP基因的鉴定[C]//植物保护21世纪展望.北京:中国科学技术出版社,1998:118-120.
[11]ARTURO R R,JORGE E.Fingerprinting of Bacillus thuringiensis type strains and isolates by using Bacillus cereus groupspecific repetitive extragenic palindromic sequence-based PCR analysis[J].Appl Environ Microbiol,2005,71(3):1346-1355.
[12]吴洪福,郭淑元.苏云金芽孢杆菌杀虫晶体蛋白结构和功能研究进展[J].东北农业大学学报,2009,40(2):118-122.
[13]TOSHIHIKO A,HIGUCHI K,MIZUKI E,et al.Nontoxic crystal protein from Bacillus thuringiensis demonstrates a remarkable structural similarity to β-pore-forming toxins[J].Proteins,2006,63(1):243-248.
[14]SOMPHOB L.Novel preparation and characterization of the α4-loop-α5 membrane-perturbing peptide from the Bacillus thuringiensis Cry4Baδ-endotoxin[J].J Biochemis Mol Biol,2006,39(3):270-277.
猜你喜欢
中国生物防治学报(2022年3期)2022-07-09
微生物学杂志(2021年2期)2021-07-01
微生物学杂志(2020年2期)2020-12-31
今日农业(2019年12期)2019-08-15
上海农业科技(2019年3期)2019-06-25
江西农业(2018年9期)2018-08-20
中国中医药信息杂志(2016年7期)2016-12-01
河北林业科技(2016年5期)2016-11-08
川北医学院学报(2015年5期)2015-12-05
中国质量与标准导报(2014年4期)2014-03-11
