
SIRT1高表达在NMDA诱导的兴奋性神经毒中的神经保护作用1)
2013-09-13 09:06:42司沛沛孙雪菲杨小荣
中西医结合心脑血管病杂志 2013年3期
司沛沛,孙雪菲,杨小荣,张 策
谷氨酸是中枢神经系统中重要的神经递质,在胚胎期神经发育、脑的兴奋性突触传递、突触可塑性方面发挥了重要作用。但是,谷氨酸过度聚集可引起细胞损伤甚至死亡,称为兴奋性神经毒,兴奋性神经毒模型是最常用、最具毒性、损伤机制最复杂、最具临床意义的一个神经损伤模型,它参与多种神经病理过程,如脑缺血损伤、各种神经退行性疾病等[1,2]。
沉默信息调节因子1(silent information regulator 1,SIRT1)是哺乳动物中第一个被发现的去乙酰化酶Sirtuin蛋白家族成员,作为一种NAD+依赖的脱乙酰基酶,它与组蛋白乙酰基酶共同维持细胞核的乙酰化平衡。SIRT1以组蛋白和多种非组蛋白作为底物,通过其去乙酰化作用调节基因转录、染色体稳定性和靶蛋白活性,进而参与能量代谢、细胞周期、细胞衰老及肿瘤发生发展等生理病理过程的调节。近年来,有关SIRT1的神经保护作用受到越来越广泛的关注。为进一步验证SIRT1可能具有的广泛的神经保护作用,本研究拟采用NMDA诱导的兴奋性神经毒细胞模型,应用(cell counting Kit-8)CCK-8分析、乳酸脱氢酶(LDH)检测、Western-blot等方法,明确SIRTI高表达在NMDA诱导的兴奋性神经毒中的保护作用。
1 材料与方法
1.1 主要试剂 SH-SY5Y人神经母细胞瘤细胞,购自上海中国科学院细胞库;胎牛血清购自Gibco公司;NMDA试剂购自sigma公司;CCK-8购自Dojindo;Lipofectamine 2000转染试剂、Opti-MEM无血清培养基购自Invitrogen公司;MEM、F12培养基,Commassie改良增强型蛋白质定量试剂盒购自武汉博士德生物工程公司;质粒提取试剂盒购自全式金生物有限公司;兔抗人SIRT1抗体购自Abcam公司;β-actin山羊抗兔二抗购自中杉金桥;Super ECL Plus超敏发光液购自普利莱基因技术有限公司。
1.2 方法
1.2.1 细胞培养 SH-SY5Y培养基为 MEM、F12,含有10%胎牛血清,100U/mL青霉素、100U/mL链霉素。将细胞以1×105的细胞浓度接种于孔板或培养瓶中,培养于37℃,5%CO2条件下,每3天以1∶2传代一次。
1.2.2 NMDA毒性诱导 采用 NMDA(500μmol/L)和甘氨酸(10μmol/L)共孵育2h,更换培养基继续培养10h,再进行后续的CCK-8、LDH测定以及蛋白提取。
1.2.3 质粒转染 按照质粒提取试剂盒说明书提取质粒后,利用脂质体介导的方法将其转入SH-SY5Y细胞中。细胞培养24 h后进行转染,质粒和lipo2000以1∶2.5混合后,加入无血清无双抗培养基中,培养6h后更换为全培养基,继续培养24 h~36h,转染效率达到最高。
1.2.4 CCK-8的测定 将培养在96孔板中的细胞在转染24h后,加入NMDA进行毒性诱导,继续培养12h,然后每孔加入CCK-8 10μL,37℃避光培养2h,上酶标仪测定,检测波长450 nm,参比波长650nm。
1.2.5 乳酸脱氢酶释放检测 将培养在六孔板中的细胞转染24h后 ,加入NMDA进行毒性诱导,继续培养12h,收集细胞培养液进行LDH活性检测。
1.2.6 Western blot检测与分析 利用Commassie法测定蛋白浓度。用10%分离胶、5%浓缩胶进行SDS-PAGE电泳,将蛋白转移到PVDF膜后,用0.3%明胶中室温封闭1h,加入一抗SIRT1(1∶5 000)、β-actin(1∶3 000)4℃孵育过夜,洗膜后加入二抗(1∶3 000)室温孵育1.5h。洗膜后加入ECL超敏发光液,曝光拍照,用分子生物学图像分析系统进行定量。β-actin条带作为内参。
1.3 统计学处理 应用SPSS13.0软件分析。数据以均数±标准差(±s)表示,采用单因素方差分析,多重比较采用LSD法。
2 结 果
2.1 SIRT1蛋白水平的改变 Western blot结果显示,与正常组相比,SIRT1过表达组中SIRT1含量上调,增加1.15倍(P<0.05);空质粒组、lipo2000组未影响SIRT1的表达水平(P>0.05)。详见图1。

图1 SIRT1表达水平的改变
2.2 SIRT1高表达对NMDA引起的细胞活力下降的作用将构建的SIRT1表达载体转入SH-SY5Y细胞中24h后,加入NMDA(500μmol/L)作用2h,继续孵育12h后采用CCK-8检测细胞活力。结果显示,而SIRT1高表达可有效抑制NMDA引起的细胞活力下降,使其活力恢复60.95%(P<0.05);与正常组相比,lipo2000、SIRT1组对细胞活力无统计学意义(P>0.05)。详见图2。
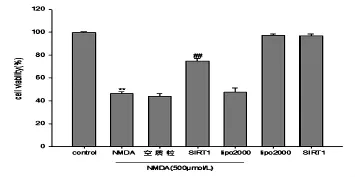
图2 SIRT1高表达对NMDA引起的细胞活力下降的观察
2.3 SIRT1高表达对NMDA引起的LDH释放的作用 在LDH实验中发现,与对照组相比,NMDA组的LDH水平增高了59.45%(P<0.05);SIRT1高表达使NMDA诱导释放的LDH减少24.26%(P<0.05);lipo2000、SIRT1组对LDH 的释放无统计学意义(P>0.05)。详见图3。
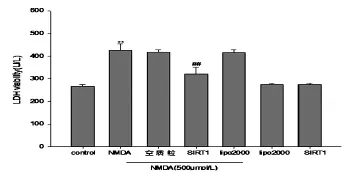
图3 SIRT1高表达对NMDA引起的LDH释放的作用
3 讨 论
谷氨酸作为中枢神经系统一种重要的神经递质,当在突出间隙中过度积聚时,可以激活突出后膜NMDA受体,使胞外Ca2+内流,激活多种蛋白酶,过度产生自由基,这些因素共同作用,引起神经元损伤甚至死亡,产生兴奋性神经毒效应[3]。
研究发现,热量限制(calorie restriction,CR)可以延长酵母及啮齿动物寿命,推迟哺乳动物年龄相关的疾病如癌症、动脉粥样硬化及糖尿病的发生,可能正是通过增加去乙酰化酶Sirtuin活性而起作用。这类蛋白修饰酶最初是在酵母中发现,被称为沉默信息调节因子2(silent information regulator 2,Sir2)。随后,人们在哺乳动物体内也发现了具有广泛去乙酰化作用的Sir2编码蛋白。在哺乳动物基因组中,目前证实存在7种Sir2同系物(SIRTs 1-7),其中SIRT1是氨基酸序列最接近Sir2的同系物。SIRT1可能通过其去乙酰化酶活性参与调节物质代谢,如抑制糖酵解、维持血糖稳定、防止脂肪变性等。最新研究表明,SIRT1高表达后通过其去乙酰化作用,激活不同信号通路可防止心肌肥大,血栓形成以及血管平滑肌的肥大。
在多种神经损伤模型,如脑缺血性损伤模型、AD大鼠模型、HD小鼠模型、肌萎缩性侧索硬化症(ALS)和脊髓和延髓肌萎缩症(SBMA)模型,通过上调SIRT1增加其去乙酰化酶活性,发挥神经保护作用。在p25转基因鼠,SIRT1过表达可以显著对抗神经退行性病;在猴的阿尔茨海默病(Alzheimer’s disease,AD)动物模型中,高表达SIRT1能减少脑内的β淀粉样蛋白(amyloid-β,Aβ),且脑内 Aβ含量与同一区域的 SIRT1含量呈负相关[4]。随后离体、在体PD模型中发现,过表达SIRT1可促进α-synuclein蛋白降解,减缓神经变性[5]。Parker和他的同事们已在线虫中证明,过表达SIRT1可以抑制HTT突变引起的神经元死亡[6];在转基因小鼠HD模型中,将SIRT1过表达可以使小鼠的运动协调能力得到显著改善,并减轻外周神经及纹状体神经元的损伤[7,8]。将SIRT1基因转染至原代神经元,发现过表达SIRT1可以拮抗由SOD1突变引起的神经毒性作用。SIRT1高表达在多种疾病损伤模型中可发挥神经保护作用。本实验中利用NMDA诱导的兴奋性神经毒模型,观察SIRT1高表达在NMDA诱导的细胞损伤中的作用。谷氨酸作为中枢神经系统一种重要的神经递质,当在突出间隙中过度积聚时,可以激活突出后膜NMDA受体,使胞外Ca2+内流,激活多种蛋白酶,过度产生自由基,这些因素共同作用,引起神经元损伤甚至死亡,产生兴奋性神经毒效应。在实验中,利用脂质体lipo2000介导的方法将质粒转入细胞中,加入NMDA诱导损伤,采用CCK-8、LDH检测发现,SIRT1高表达可以拮抗NMDA神经毒性损伤,表现为细胞活力增加,LDH释放减少。
而上述保护效应由SIRT1对底物P5 3、FOXOs、Ku7 0、PGC-1α、TAFI68、P300、PCA 等 的 去 乙 酰 化 作 用 介 导 的[9,10]。相应实验观察显示,SIRT1可以通过使P53去乙酰化,从而负性调节其活性,使细胞免于凋亡,继续分裂生长;SIRT1通过使凋亡激活蛋白FOXOs去乙酰化而抑制其转录,使神经元免于凋亡[11];SIRT1可以使NF-κB亚基p65的赖氨酸310位点去乙酰化而降低其活性,防止细胞损伤;SIRT1通过使HSF1(heat shock factor 1,HSF1)去乙酰化,促进HSF1与Hsp70启动子结合,激活 Hsp70,减少细胞凋亡[12]。
本实验表明,在NMDA引起的神经毒过程中,SIRT1作用可能受到抑制,介导神经损伤,SIRT1高表达证实了可以拮抗NMDA引起的细胞损伤作用,发挥神经保护作用。但是,SIRT1具体的神经保护机制尚未明确,需要进一步研究证实。
[1]Gc P.Neuroprotection by NMDA receptor antagonists in a variety of neuropathologies[J].Current Drug Targets,2001,2:241-271.
[2]Robert J.Dempsey MD.Attenuation of brain edema,blood-brain barrier breakdown,and injury volume by ifenprodil,apolyaminesite N-Methyl-D-aspartate receptor antagonist,after experimental traumatic brain injury in rats[J].Neurosurgery,2000,47(2):399-404.
[3]Hynd MR,Scott HL,Dodd PR.Glutamate-mediated excitotoxicity and neurodegeneration in Alzheimer’s disease[J].Neurochem Int,2004,45(5):583-595.
[4]Qin W,Chachich,M,Lane,M,etal.Calorie restriction attenuates Alzheimer’s disease type brain amyloidosis in Squirrel monkeys(Saimiri sciureus)[J].J Alzheimers Dis,2006,10(4):417-422.
[5]Wareski P,Vaarmann A,Choubey V,etal.PGC-1{alpha}and PGC-1{beta}regulate mitochondrial density in neurons[J].J Biol Chem,2009,284(32):21379-21385.
[6]Parker JA,Arango M,Abderrahmane S,etal.Resveratrol rescues mutant polyglutamine cytotoxicity in nematode and mammalianneurons[J].Nat Genet,2005,37(4):349-350.
[7]Ho DJ,Calingasan NY,Wille E,etal.Resveratrol protects against peripheral deficits in a mouse model of Huntington’s disease[J].Exp Neurol,2010,225(1):74-84.
[8]Jeong H,Cohen DE,Cui L,etal.Sirt1mediates neuroprotection from mutant huntingtin by activation of the TORC1and CREB transcriptional pathway[J].Nat Med,2012,18:159-165.
[9]Kim D,Nguyen MD,Dobbin MM,etal.SIRT1deacetylase protects against neurodegeneration in models for Alzheimer’s disease and amyotrophic lateral sclerosis[J].EMBO J,2007,26(13):3169-3179.
[10]Han S,Choi JR,Soon Shin K,etal.Resveratrol upregulated heat shock proteins and extended the survival of G93A-SOD1mice[J].Brain Res,2012,1483:112-117.
[11]Anekonda TS,Reddy PH.Neuronal protection by sirtuins in Alzheimer’s disease[J].J Neurochem,2006,96(2):305-313.
[12]Donmez G,Arun A,Chung CY,etal.SIRT1protects against alpha-synuclein aggregation by activating molecular chaperones[J].J Neurosci,2012,32(1):124-132.
猜你喜欢
浙江大学学报(医学版)(2023年6期)2023-05-11 11:09:30
中国生物化学与分子生物学报(2022年8期)2022-09-08 00:39:50
中西医结合心血管病电子杂志(2018年28期)2018-11-19 11:04:54
食品科学(2018年10期)2018-05-23 01:27:28
中国组织化学与细胞化学杂志(2016年4期)2016-02-27 11:15:53
中国现代医学杂志(2015年26期)2015-12-23 11:04:20
西南医科大学学报(2015年1期)2015-08-22 13:01:46
江苏农业科学(2015年1期)2015-04-17 23:53:02
中国当代医药(2015年16期)2015-03-01 02:03:13
中国当代医药(2015年9期)2015-03-01 02:01:59
