
环境和扩散对草地群落构建的影响
2013-09-07 02:53王孝安王世雄郑维娜刘史力
生态学报 2013年14期
王 丹,王孝安,郭 华,王世雄,郑维娜,刘史力
(陕西师范大学生命科学学院,西安 710062)
如何解释人们观察到的物种时空分布模式一直是群落生态学的核心内容之一。在这个领域内,最关键的问题是物种共存机制[1-5]。过去人们一直在生态位理论和中性理论间争论不休。生态位理论认为每个物种都有自身最为适宜的生境,物种根据自己的存活能力,在不同的资源条件下进行权衡[6-7],物种的特性是其对物理和生物环境进化适应的一种体现[8-9],因而环境限制是生态位理论的一个重要方面。而中性理论假定所有个体在同一营养级内是功能等价的[4,10-12],强调扩散和随机作用的重要性[13],扩散限制的空间表现即为地理距离。但近年来,越来越多的生态学家则认为物种共存机制应是二者的整合[14]。
对于上述观点,不同的研究得到了不同的结果。Hanna等[15]在对亚马逊西部森林的研究中指出,环境变量能更多的解释不同样地之间植物群落组成的差异;空间自相关的随机、斑块和环境的共同作用对物种的分布和组成也有很好的诠释。Benjamin等[16]研究表明对于物种多度和物种分布的影响,生态位理论占主导地位,而中性理论几乎没有得到支持。Turnbull[17]通过对白垩草地播种试验的研究表明,在先锋物种定居的过程中,不同大小的种子通过占有不同的生态位而达到共存,即使到了成年阶段,种间也不存在明显的竞争。Jaana等[18]从植物的多样性和优势种入手,研究了亚马逊西部植物的共存机制,发现扩散和环境对物种的组成都有影响,但扩散作用更为显著。
类似的研究还有很多,而得到的结果并不一致。且大部分试验都集中在物种丰富的热带雨林地区,对于群落组成相对简单,气候条件相对苛刻的黄土高原来说,这样的研究还没有报道。鉴于此,本文以黄土高原子午岭地区3种草地群落(5a的弃耕地、阴坡和阳坡的草地)为例,拟通过植物群落的野外调查分析,探讨扩散和环境对该地区群落构建的影响,以期为这一问题的解决提供理论依据。
1 材料和方法
1.1 研究区域概况
马栏林区位于陕西省旬邑县的东北部,地处黄土高原中部。地理位置为108°27'—108°52'E,35°9'—35°33'N,东西宽约40 km,南北长约43 km,海拔为1 200—1 700 m,坡度约为25°。该地区的土壤类型为成土母质的风积黄土,结构疏松,机械组成多为中壤,富含钙质,土层深约50 cm,pH值在7—9范围。该区的气候特点属于暖温带半湿润地区,据气象站记载,这里全年平均气温在7.4—8.5℃,最低气温为-27.7℃,最高气温为36.7℃。年平均降雨量为580 mm,多集中在7月、8月和9月,干旱季节为12月、1月和2月,光照充足,湿热同期,益于林木生长。但是降水季节分布不均,容易出现春旱和伏旱,影响林木种子的萌发和造林成活率,对森林更新有一定影响[19]。
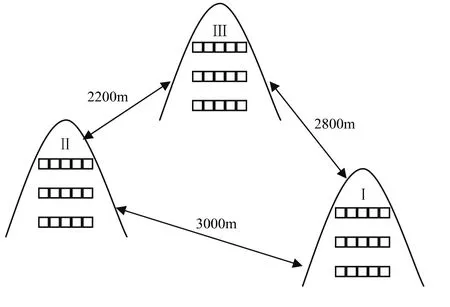
图1 样地空间分布模拟图Fig.1 Simulation diagram of spatial distribution of sampling sets
该地区的植被类型复杂多样,有次生的辽东栎(Quercus wutaishanica)林、油松(Pinus tabulaeformis)林、油松+辽东栎混交林,和少量的次生山杨(Populus davidiana)林、白桦(Betula platyphylla)林以及二者与辽东栎的混交林、人工油松林、人工刺槐(Robinia pseudoacacia)林和灌丛、草地、退耕地等。
1.2 试验设计和样本采集
在对子午岭植被类型进行全面考察的基础上,选择3块完全由草本构成的山坡作为研究样地(图1),分别编号为Ⅰ、Ⅱ、Ⅲ,其面积分别为 1.2、1.5、1.5 hm2。沿着与坡向垂直的方向,在每块山坡的上坡位、中坡位和下坡位分别设置长5 m、宽1 m的样带,即每条样带包含5个1 m×1 m的样方。在每条样带中记录物种种类、数量、高度和盖度数量指标,同时记录样地的经纬度、每个样地中样带的坡度、坡向、坡位和海拔等环境指标,以及所有样地九条样带中任意两条样带之间的距离。在每条样带中随机选择3点,用Hydra土壤水分/盐分测定仪分别测定每条样带中这3点的土壤温度、湿度、电解质和电导率,待该点土壤指标测完后,用土钻在该点进行20 cm深度的取土,将3个样方的土壤混匀后装袋,低温保存,带回实验室阴干,去除石砾、落叶等杂物,研磨后过筛。按照鲍士旦[20]的土壤处理方法,用SYSTEA公司生产的第三代连续流动分析仪(FlowsysⅢ)测定速效氮含量。
统计每个样地物种的重要值,剔除重要值小于0.05的物种,建立植物群落数据库(表1),重要值算法如下:
草本的重要值 =(相对密度+相对盖度+相对频度)/300

表1 3个样地中重要值大于0.05的物种组成Table1 Species whose importance value is more than 0.05 in the three sites
1.3 数据分析方法
在坡向定值时,参考王孝安[21]的转换方法,其数值越大,表示越向阳、越干热。样地的坡向、坡度、海拔、温度、湿度、电解质、电导率和速效氮构成环境因子矩阵。Mantel test用于分析物种相似矩阵与环境矩阵、物种相似矩阵与距离矩阵、距离矩阵与环境矩阵之间的相关程度,其中物种相似性选择Bray-Curtis指标,环境矩阵和空间距离矩阵都转化为欧氏距离矩阵。主轴邻距法利用空间变量结合环境因子,通过方差分解方法,分析各变量对物种组成的相对影响[22]。典范对应分析(CCA)用于分析每一个环境因子对物种组成的影响。以上分析分别在 R-2.15.1软件中完成。
2 结果
2.1 环境和扩散与草地群落构建的相关性
由Mantel test分析可以得到如下结果:空间地理距离与群落组成相似性的相关值为0.7542,P值为0.0026;而纯粹的地理距离与群落组成相似性的相关值为0.6531,P值为0.0048。环境距离与群落组成相似性的相关值为0.5554,P值为0.0100;而纯粹的环境距离与群落组成相似性的相关值为0.4162,P值为0.0124。由此可以看出样方间群落组成的相似性与地理距离和环境距离都显著相关,且与空间地理距离的相关程度高于环境距离(表2)。
2.2 环境和扩散对草地群落构建的解释量
为探究环境变量和空间地理距离对群落构建的影响,通过主轴邻距法将环境因子和空间距离进行方差分解,分析各变量对物种组成的独立贡献率。首先对环境因子进行前向选择,结果表明速效氮和土壤湿度对物种组成的影响是显著。为了进一步了解各个环境因子的作用,将其进行CCA排序,通过CCA排序结果表明土壤湿度和速效氮对物种分布有影响,尤其是速效氮(r2=0.8178,P=0.01)的作用较为突出(表3)。
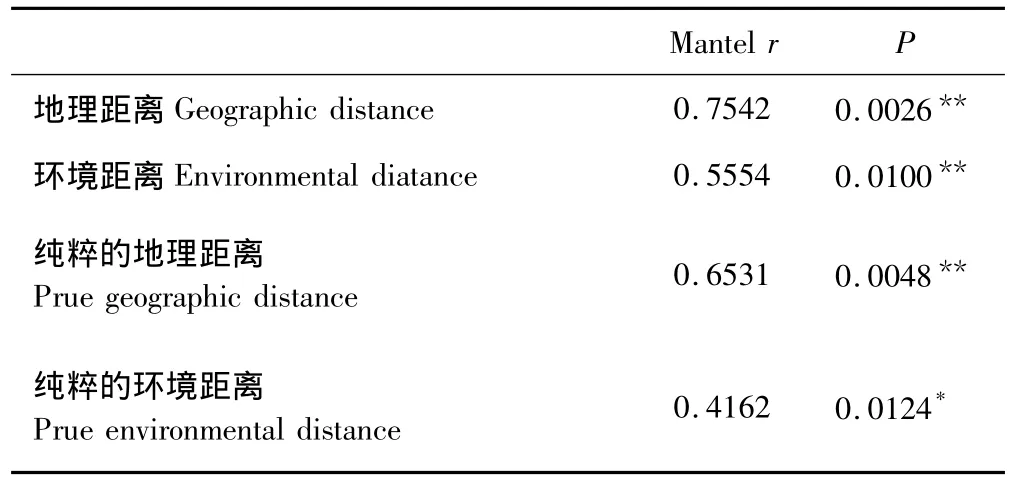
表2 群落组成的相似性与地理距离和环境距离之间的相关性Table2 Correlations between similarity in community composition,geographic distance and environment distance
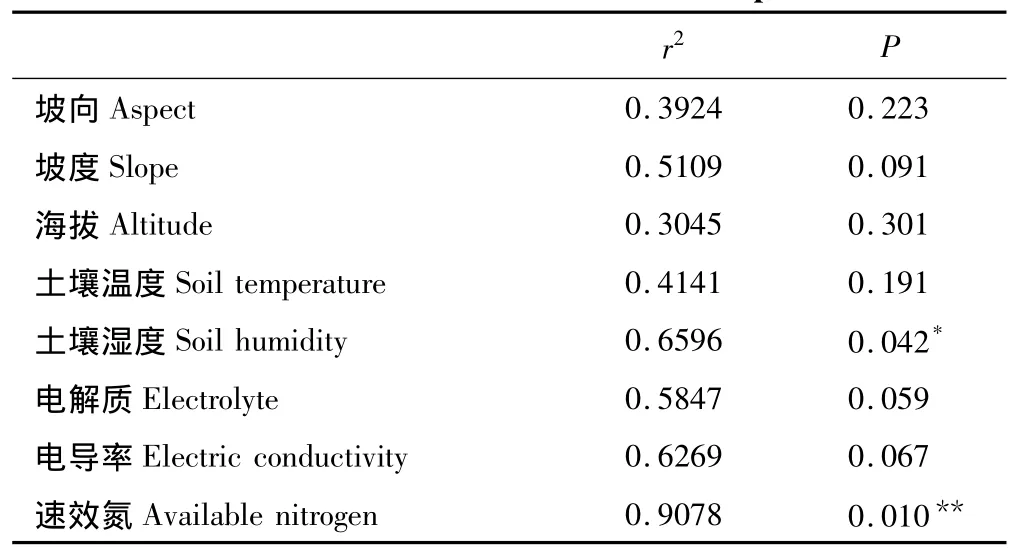
表3 环境因子对物种分布的影响Table3 The influence of environmental factors on species distribution
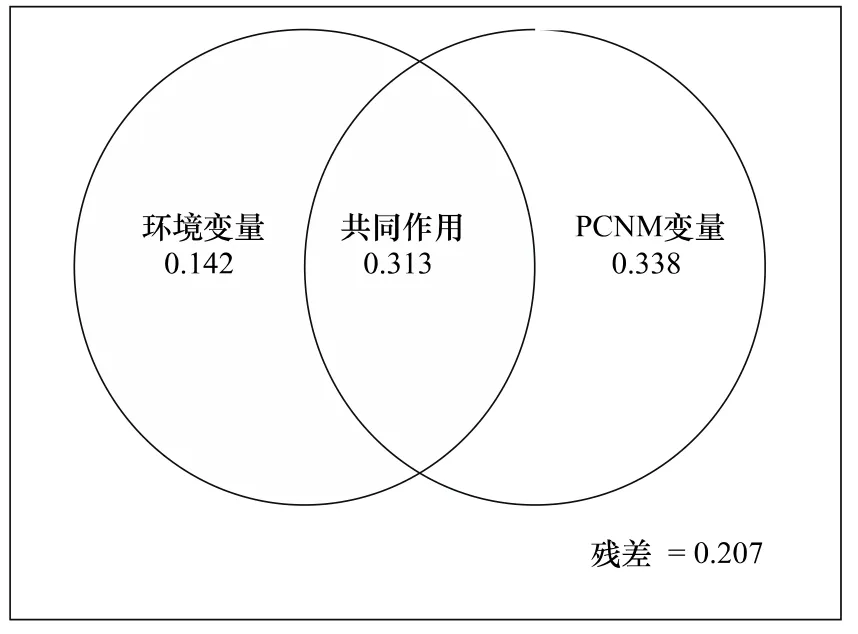
图2 方差分解图Fig.2 Variation partitioning results
方差分析结果如图2所示。环境变量和PCNM变量二者共同解释了群落构建的79.3%,而未被解释的为20.7%,其分别独立解释量为14.2%和33.8%,而和彼此配合后的解释量为45.5%和65.1%,说明PCNM变量的影响大于环境变量的影响,且环境变量与之协同作用。即物种组成受空间距离的影响大于环境的影响。
2.3 环境和扩散对草地群落构建的相对影响
地理距离和环境因子合力解释了物种组成的79.3%(图2)。纯粹的地理距离解释了绝大部分物种组成,其值为33.8%,而纯粹的环境因子解释的物种组成为14.2%,前者是后者的2倍多(图2)。由曲线相关图分析可知:整体而言,样方间植物相似性随地理距离的增加而下降,但在地理距离大约为2800 m的C点和3000 m的D点时,植物相似性高于地理距离约为2200 m的B点(图3);而在此处C点和D点的环境相似性值也成反弹趋势,高于B点(图4)。说明环境变量解释了植物组成相似性的部分原因。而随着环境差异性的增大,植物相似性表现为下降趋势(图5)。
3 讨论
生态学的核心是群落生态学,而群落生态学的核心是物种共存。长期以来,人们对于这一问题的研究和争论从未停止,而结论尚未达成一致。自20世纪60年代以来,生态位理论已在种间关系、群落结构和生物多样性中得到了广泛的应用[23]。在本研究中环境变量对植物相似性的解释量为45.5%(图2),表明生态位对群落构建有一定的作用,但是剔除空间距离后,其解释量为14.2%(图2),作用比较小。而之前在很多表明生态位占主导作用的研究中,都提到土壤类型在决定物种分布时扮演了重要的角色[24-30]。而子午岭地区土壤类型较一致,均为风积黄土,各样地间差异不明显,反而是速效氮在所有的环境因子中的作用最为显著。这与Tilman[31]的研究结果一致,其在1984年对草地群落进行研究表明:在草地群落中,土壤中的氮元素是主要的限制资源。在本研究中土壤湿度对植物相似性也有一定的影响,由于阳坡较阴坡日照时间长,土壤温度高,所以土壤湿度相对较低,使得处于半干旱区的黄土高原的植被组成,对于土壤湿度表现出一定的响应。整体而言随着地理距离的增加,环境相似性表现为下降趋势(图4),存在较为明显的环境梯度,即存在生态位的分化;但环境变量对植物相似性的解释量很小,说明生态位理论对该地区植物群落的构建作用不显著。
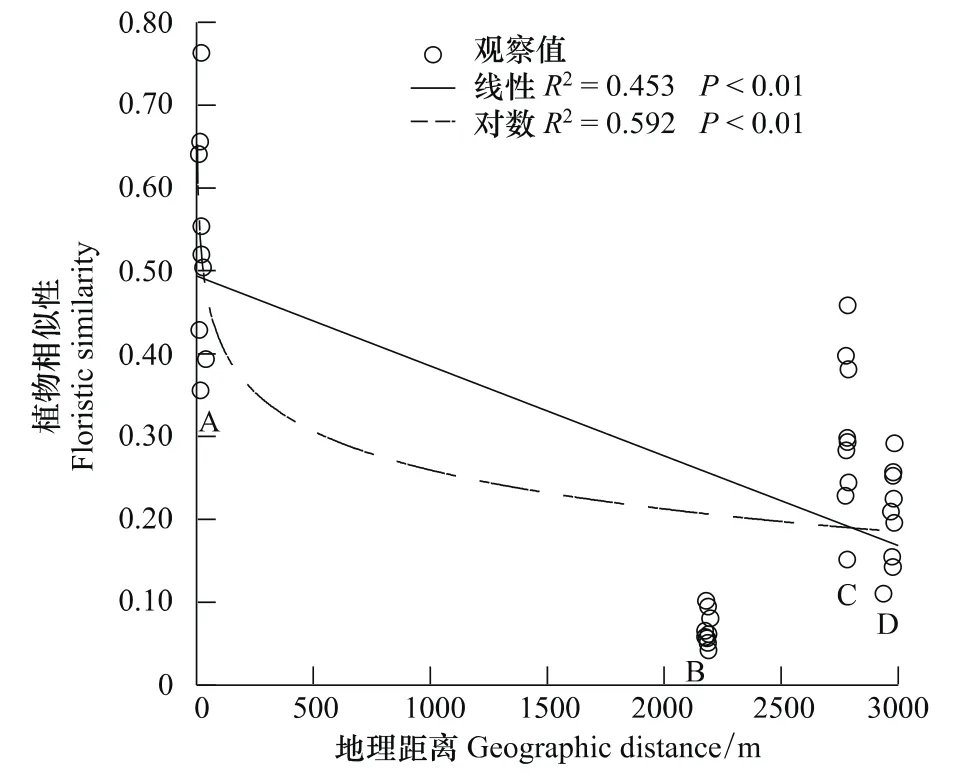
图3 植物相似性与地理距离间的曲线相关图Fig.3 The relevant figure curve between floristic similarity and geographic distance
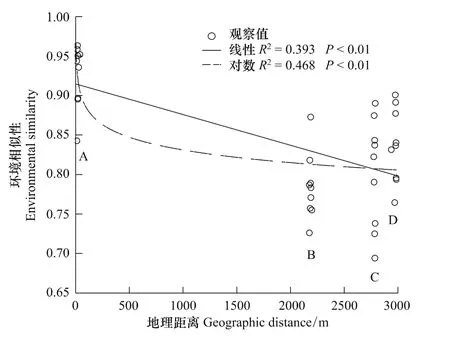
图4 环境相似性与地理距离间的曲线相关图Fig.4 The relevant figure curve between environmental similarity and geographic distance
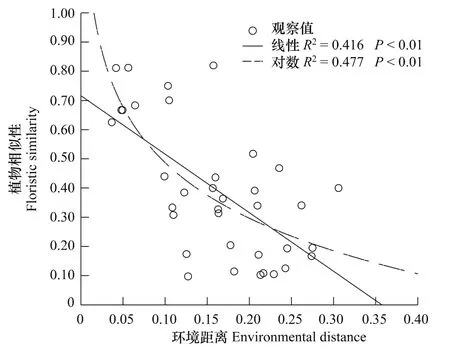
图5 植物相似性与环境距离的曲线相关图Fig.5 The relevant figure curve between floristic similarity and environmental distance
中性理论认为物种组成是物种入侵和灭绝的结果,空间距离制约了种子的扩散,从而导致随着地理距离的增加,样方间植物的相似性下降[4,32-33]。本研究中地理距离和环境距离共同解释了植物相似性的79.3%(图2),剔除环境因子的影响,地理距离解释了植物相似性的33.8%(图2);即去除空间解释的群落组成相似性中由环境解释的部分后,其解释组分仍然较大,这提供了对中性理论的支持[34],随着地理距离的增加,植物相似性表现为明显的下降趋势(图3),说明距离限制的确是该地区植物构建过程中的限制因素;从而很好地印证了中性理论在该地区植物群落构建中的作用。这与Chave[13]以及 Condit[33]等以热带雨林为研究目标所得到的结果一致。一方面由于黄土高原经受流水侵蚀,形成沟壑纵横,墚、峁广布的破碎地貌,因而在不同的时间尺度上影响着基因、个体、种群和物种的运动[35-36];另一方面,黄土高原气候脆弱,降雨年际变化大,且年内不均[19],使多数树木不落叶,枯枝落叶层难以形成,甲虫、蜗牛等因缺乏栖息地而减少,近而影响物种扩散[37]。此外本文选取的草地群落处于演替前期,而演替前期的物种繁殖力较高,传播距离相对较远,可先定居环境[38],所以会得到中性理论扮演了更为重要的角色的结果。
中性理论与生态位理论在群落构建过程中的作用一直存在争议,在本研究中二者对子午岭地区草地群落构建都起作用,虽然大量的数据表明中性理论占据着主导地位,但不可否认生态位理论对其的影响。近年来,研究者倾向于认为生态位分化与中性理论对群落的构建都有作用,源于二者的相对贡献与研究尺度和生态系统类型有关[39]。因而提出将生态位理论与中性理论整合,同时发展包含随机的生态位模型或者近中性模型。正如Chase[40]所提到的,如能将中性理论和生态位理论的关键要素结合起来解释群落多样性模式,将会是一次真正的生态学突破。
4 结论
为了探讨植物群落的构建机制,本文以黄土高原子午岭地区草地植物群落为例,研究了样方间植物相似性与空间地理距离和环境差异之间的关系,比较距离限制的中性理论和环境资源分化的生态位理论二者的相对贡献。结果表明:扩散和环境对黄土高原草地群落构建都有影响,但扩散作用占据主导地位。
[1] Tokeshi M.Niche apportionment or random assortment:species abundance patterns revisited.The Journal of Animal Ecology,1990,59(3):1129-1146.
[2] Tilman D,Pacala S W.The maintenance of species richness in plant communities//Rickleffs R E,Schluter D,eds.Species Diversity in Ecological Communities.Chicago:Chicago University Press,1993:13-25.
[3] Chesson P.Mechanisms of maintenance of species diversity.Annual Review of Ecology and Systematics,2000,31(1):343-366.
[4] Hubbell SP.The Unified Neutral Theory of Biodiversity and Biogeography.Princeton:Princeton University Press,2001.
[5] Zhou SR,Zhang D Y.Neutral theory in community ecology.Chinese Journal of Plant Ecology,2006,30(5):868-877.
[6] Chesson P L,Warner R R.Environmental variability promotes coexistence in lottery competitive systems.The American Naturalist,1981,117(6):923-943.
[7] Chesson P L.Coexistence of competitors in spatially and temporally varying environments:a look at the combined effects of different sorts of variability.Theoretical Population Biology,1985,653(28):263-287.
[8] Ackerly D D,Dudley SA,Stultan SE,Schmitt J,Coleman JS,Linder CR,Sandquist D R,Geber M A,Evans A S,Dawson T E,Lechowicz M J.The evolution of plant ecophysiological traits:recent advances and future directions.BioScience,2000,50(11):979-995.
[9] Gillespie R.Community assembly through adaptive radiation in Hawaiian spiders.Science,2004,303(5656):356-359.
[10] Bell G.The distribution of abundance in neutral communities.The American Naturalist,2000,155(5):606-617.
[11] Caswell H.Community structure:a neutral model analysis.Ecological Monographs,1976,46(3):327-345.
[12] Hubbell SP.Neutral theory in community ecology and the hypothesis of functional equivalence.Functional Ecology,2005,19(1):166-172.
[13] Chave J.Neutral theory and community ecology.Ecology Letters,2004,7(3):241-253.
[14] Niu K C,Liu Y N,Shen Z H,He F L,Fang JY.Community assembly:the relative importance of neutral theory and niche theory.Biodiversity Science,2009,17(6):579-593.
[15] Tuomisto H,Ruokolainen K,Yli-Halla M.Dispersal,environment,and floristic variation of western Amazonian forests.Science,2003,299(5604):241-244.
[16] Gilbert B,Lechowicz M J.Neutrality,niches,and dispersal in a temperate forest understory.Proceedings of the National Academy of Sciences of the United States of America,2004,101(20):7651-7656.
[17] Turnbull L A,Manley L,Rees M.Niches,rather than neutrality,structure a grassland pioneer guild.Proceedings of the Royal Society B:Biological Sciences,2005,272(1570):1357-1364.
[18] Vormisto J,Svenning J C,Hall P,Balslev H.Diversity and dominance in palm(Arecaceae)communities in terra firme forests in the western Amazon basin.Journal of Ecology,2004,92(4):577-588.
[19] Department of Forestry of Shaanxi Province.Manual of Shaanxi Forestry.Beijing:China Forestry Press,1964:22-53.
[20] Bao SD.Soil Agricultural Chemical Analysis.Beijing:China Agriculture Press,2003:47-60.
[21] Wang X A.Multivariate analysis and environmental interpretation of plant communities in Maqu,South Gansu.Acta Ecologica Sinica,1997,17(1):61-65.
[22] Borcard D,Legendre P.All-scale spatial analysis of ecological data by means of principal coordinates of neighbour matrices.Ecological Modelling,2002,153(1/2):51-68.
[23] Wang G.A discussion on some aspects of niche theory.Journal of Lanzhou University:Natural Science Edition,1990,26(2):109-113.
[24] Clark D B,Clark D A,Read JM.Edaphic variation and the mesoscale distribution of tree species in a neotropical rain forest.Journal of Ecology,1998,86(1):101-112.
[25] Clark D B,Palmer M W,Clark D A.Edaphic factors and the landscape-scale distributions of tropical rain forest trees.Ecology,1999,80(8):2662-2675.
[26] Gartlan J S,Newbery D M C,Thomas D W,Waterman PG.The influence of topography and soil phosphorus on the vegetation of Korup Forest Reserve,Cameroun.Plant Ecology,1986,65(3):131-148.
[27] John R,Dalling J W,Harms K E,Yavitt J B,Stallard R F,Mirabello M,Hubbell SP,Valencia R,Navarrete H,Vallejo M,Foster R B.Soil nutrients influence spatial distributions of tropical tree species.Proceedings of the National Academy of Sciences of the United States of America,2007,104(3):864-869.
[28] Newbery D M C,Gartlan JS,Mckey D B,Waterman P G.The influence of drainage and soil-phosphorus on the vegetation of Douala-Edea Forest Reserve,Cameroun.Plant Ecology,1986,65(3):149-162.
[29] Pélissier R,Dray S,Sabatier D.Within-plot relationships between tree species occurrences and hydrological soil constraints:an example in French Guiana investigated through canonical correlation analysis.Plant Ecology,2002,162(2):143-156.
[30] Poulsen A D,Tuomisto H,Balslev H.Edaphic and floristic variation within a1-ha plot of lowland Amazonian rain forest.Biotropica,2006,38(4):468-478.
[31] Tilman G D.Plant dominance along an experimental nutrient gradient.Ecology,1984,65(5):1445-1453.
[32] Chave J,Leigh E G Jr.A spatially explicit neutral model of β-diversity in tropical forests.Theoretical Population Biology,2002,62(2):153-168.
[33] Condit R,Pitman N,Leigh E GJr,Chave J,Terborgh J,Foster R B,Núñez P,Aguilar S,Valencia R,Villa G,Muller-Landau H C,Losos E,Hubbell SP.Beta-diversity in tropical forest trees.Science,2002,295(5555):666-669.
[34] LalibertéE.Analyzing or explaining beta diversity?.Comment.Ecology,2008,89(11):3232-3237.
[35] Minor E S,Urban DL.A graph-theory framework for evaluating landscapeconnectivity and conservation planning.Conservation Biology,2008,22(2):297-307.
[36] Clergeau P,Burel F.The role of spatio-temporal patch connectivity at the landscape level:an example in a bird distribution.Landscape and Urban Planning,1997,38(1/2):37-43.
[37] Steinitz O,Heller J,Tsoar A,Rotem D,Kadmon R.Environment,dispersal and patterns of species similarity.Journal of Biogeography,2006,33(6):1044-1054.
[38] Rees M,Condit R,Crawley M,Pacala S,Tliman D.Long-term studies of vegetation dynamics.Science,2001,293(27):650-655.
[39] Legendre P,Mi X C,Ren H B,Ma K P,Yu M J,Sun Y F,He F L.Partitioning beta diversity in a subtropical broadleaved forest of China.Ecology,2009,90(3):663-674.
[40] Chase J M.Towards a really unified theory for metacommunities.Functional Ecology,2005,19(1):182-186.
参考文献:
[5] 周淑荣,张大勇.群落生态学的中性理论.植物生态学报,2006,30(5):868-877.
[14] 牛克昌,刘怿宁,沈泽昊,何芳良,方精云.群落构建的中性理论和生态位理论.生物多样性,2009,17(6):579-593.
[19] 陕西省林业厅.陕西林业手册.北京:中国林业出版社,1946:22-53.
[20] 鲍士旦.土壤农化分析.北京:中国农业出版社,2003:47-60.
[21] 王孝安.甘南玛曲植物群落的多元分析与环境解释.生态学报,1997,17(1):61-65.
[23] 王刚.生态位理论若干问题探讨.兰州大学学报:自然科学版,1990,26(2):109-113.
猜你喜欢
数学物理学报(2022年5期)2022-10-09
昆明医科大学学报(2022年2期)2022-03-29
食品安全导刊(2021年20期)2021-08-30
河北画报(2020年8期)2020-10-27
英语学习(上半月)(2019年9期)2019-10-10
米娜·女性大世界(2016年8期)2016-08-17
浙江大学学报(工学版)(2016年2期)2016-06-05
工业设计(2016年11期)2016-04-16
水生生物学报(2015年1期)2015-02-28
食品工业科技(2014年11期)2014-03-11
