
CaCl2处理对草莓采后品质及灰霉病害的影响
2013-09-04 10:13王若兰宋永令
食品工业科技 2013年13期
赵 妍,杨 超,王若兰,宋永令
(河南工业大学粮油食品学院,河南郑州450001)
果蔬是人类不可或缺的食物资源,因其含有丰富的营养物质而易遭受病害威胁。果蔬的采后病害以真菌性病害最为严重,病原菌侵染果实,同时可以分泌次级代谢产物,常常引发食品安全问题[1]。钙盐作为一种品质保护剂,常用于果蔬采后贮藏,对乙烯的生成、呼吸速率和果实酶的活性有诱导作用[2]。草莓作为典型的非呼吸性跃变型果实,耐贮藏性较差,果皮薄,果肉含糖量较高,较易发生采后病害。灰霉病是草莓的常见真菌性病害。本研究探讨了CaCl2处理对草莓采后贮藏品质、灰霉病害及果实抗病相关物质代谢的影响,为其在草莓采后贮藏保鲜方面的应用提供理论依据。
1 材料与方法
1.1 材料与仪器
试验用草莓品种为丰香(Toyonoka),采摘自郑州市中牟草莓生产基地,采摘后即运回实验室。选取成熟度相同且大小一致、无机械损伤与病虫害痕迹的草莓为供试果实。
SYQ-DSX-280B型手提式不锈钢蒸汽灭菌锅上海申安医疗器械厂;HW型智能恒温恒湿箱 宁波东南仪器有限公司;752型紫外可见分光光度计 上海菁华科技仪器有限公司;GL-20G-Ⅱ型高速冷冻离心机 上海安亭科学仪器厂;101-2型电热鼓风恒温干燥箱 上海浦鸿仪器厂;SW-CJ-1D型单人净化工作台 苏州净化设备有限公司;PHS-3C型酸度计 上海大普仪器厂;2W型折射仪 上海易测仪器有限公司;HH-S型恒温水浴锅 巩义市英峪予华仪器厂;PL203电子天平 梅特勒-托利多仪器上海有限公司;CX21型显微镜 北京京瑞天下科技有限公司。
病原菌灰葡萄孢菌(Botrytis cinerea)分离于自然发病的草莓果实,保藏于马铃薯葡萄糖琼脂(PDA)培养基。使用前,病原菌活化培养7d,之后用含有0.05%吐温-20的无菌水冲洗孢子。使用血球计数板计数,并用无菌水调整至浓度为5×104个/mL的孢子悬浮液,备用。
1.2 草莓果实预处理
草莓果实经1%次氯酸钠溶液消毒1min后,分别浸入浓度为1%、2%、5%和10%的CaCl2溶液中,浸泡10min后取出沥干,以无菌水浸泡组为对照。
1.3 果实主要品质指标测定
草莓经预处理后,将果实置于恒温恒湿箱中10℃条件下贮藏4d。每组20个果实,试验重复三次。每天取样测定果实主要品质指标。维生素C含量采用国标GB/T 5009.86-2003测定。可滴定酸值采用国标GB 12293-1990电位滴定法测定。可溶性固形物含量采用国标GB12295-1990折射仪法测定。
1.4 草莓病害相关指标测定
1.4.1 灰霉菌菌落直径 将灰葡萄孢孢子悬浮液涂布于PDA培养基上培养5d,制成备用平板。配制10倍于待测浓度的CaCl2溶液,与新配制的PDA培养基以1∶9比例混合均匀,倒皿成含药平板。之后用打孔器在备用平板边缘制取直径为5mm的菌饼,挑取菌饼放入含药平板中央,长气生菌丝的一面朝下。以不含CaCl2的平板为对照。将平板放入27℃恒温培养箱中培养5d。采用十字交叉法测量菌落直径,每种浓度重复三次。
1.4.2 孢子萌发率 用CaCl2配制PDB培养基(无琼脂PDA),使培养基中CaCl2浓度分别达到试验所需浓度(1%、2%、5%、10%),用接种环挑取灰葡萄孢置于PDB培养基中,25℃下恒温震荡培养15h,用无菌滴管吸取培养液滴于载玻片上镜检并计算孢子萌发率。每种CaCl2浓度下的试验重复三次。
1.4.3 果实发病率 经预处理草莓果实于赤道部位打孔(3mm ×3mm),注入浓度为5×104个/mL的灰葡萄孢孢子悬浮液15μL,之后果实贮藏于4℃。15d后测定果实发病率,每种CaCl2处理浓度选取30个果实,试验重复三次。
1.5 诱导抗病性相关物质的测定
草莓果实经预处理后,贮藏于4℃,定期取样测定木质素含量和苯丙氨酸解氨酶(PAL)、过氧化氢酶(CAT)、过氧化物酶(POD)活性。
1.5.1 木质素含量测定 参照Rodrigues[3]方法测定并作部分修改。称取果肉组织1g,用95%乙醇进行研磨,经离心后抽滤,并用乙醇和乙醇-正己烷冲洗,将抽滤之后所得草莓残渣放入铝盒,105℃干燥1h。干燥后放入10mL具塞试管中,加入3mL乙酰溴-乙酸(体积比1∶3)的混合试剂,70℃下水浴30min,取出放入冰水中,向试管中加入0.9mL 2mol/L的NaOH以终止反应。用冰乙酸定容至10mL,再经离心后取上清液0.5mL于10mL具塞试管中定容,混匀后于280nm处测定其吸光度。结果用吸光度表示。
1.5.2 PAL、CAT和 POD活性测定 参考曹建康等[4]编写的《果蔬采后生理生化实验指导》。
1.6 数据统计与分析
采用SAS数据分析软件对数据进行邓肯检验,结果以均值±标准偏差表示(n=3)。
2 结果与分析
2.1 CaCl2处理对草莓果实贮藏品质的影响
由于呼吸作用和维生素C自身的氧化作用有关,采后草莓维生素C含量随贮藏期延长而逐渐下降(图1A)。CaCl2处理对果实维生素C有一定的保持能力,且与处理浓度有关。在整个贮藏期内,经由1%和2%浓度CaCl2处理的果实其维生素C含量与对照组无显著差异;而当CaCl2浓度为5%和10%时,处理组果实维生素C含量显著高于对照组(p<0.05),贮藏4d后,其含量分别为对照组的1.47倍和1.64倍。说明适宜浓度的CaCl2处理具有提高草莓果实维生素C含量的作用。
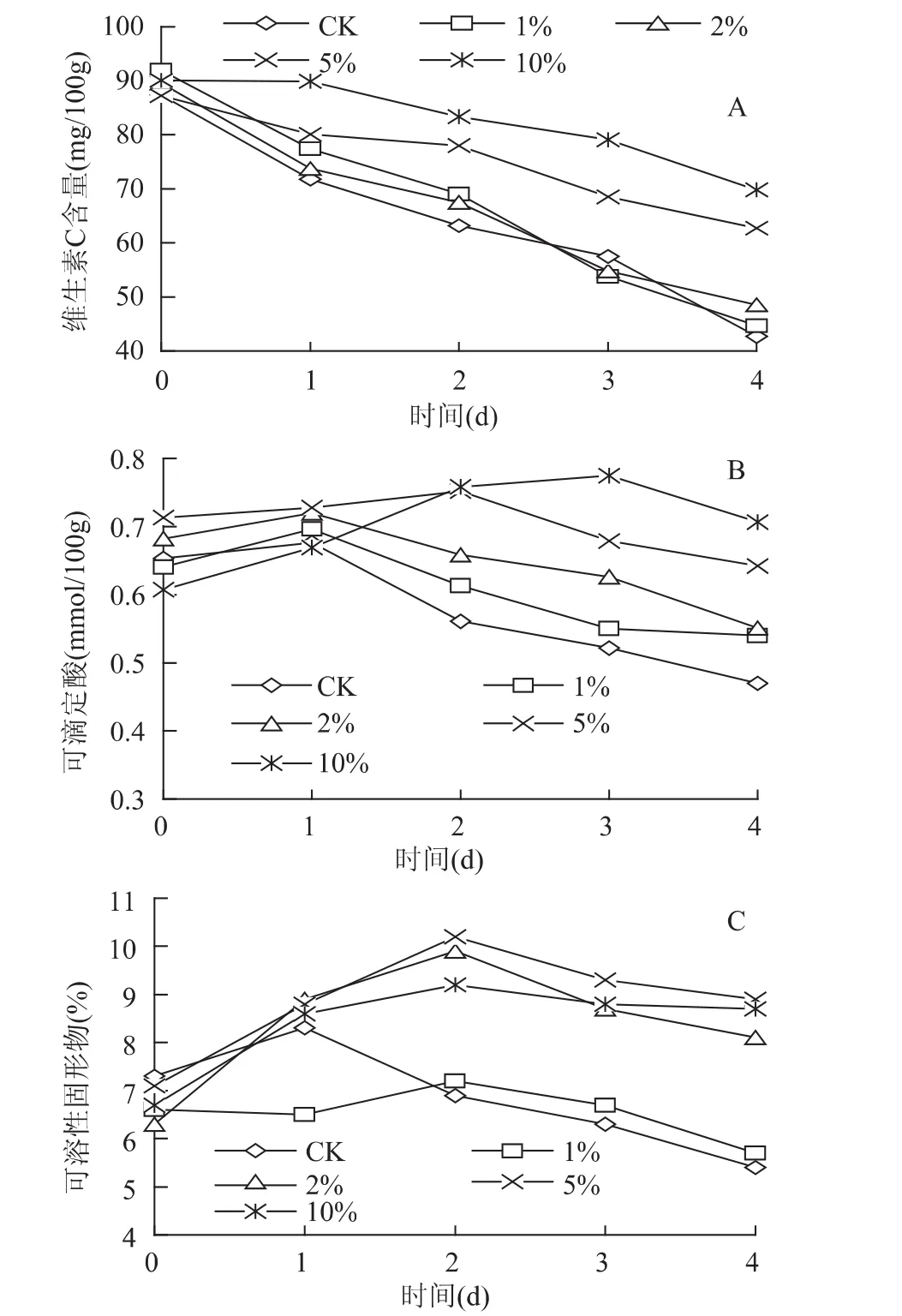
图1 氯化钙浸泡处理对草莓果实(A)维生素C、(B)可滴定酸及(C)可溶性固形物含量的影响Fig.1 Influence of CaCl2soaking on the(A)vitamin C,(B)titratable acidity and(C)soluble solids contents of strawberry fruit
可滴定酸和可溶性固形物是草莓果实品质的重要指标。如图1B所示,贮藏期间,处理组与对照组草莓果实的可滴定酸值均呈先上升后下降的趋势。对照组及低浓度CaCl2处理组(1%和2%)果实可滴定酸值在贮藏1d后即开始迅速下降;而高浓度CaCl2处理组(5%和10%)果实可滴定酸值分别在贮藏2和3d后开始下降。说明高浓度CaCl2处理具有延缓草莓果实可滴定酸含量下降的作用。
同可滴定酸类似,贮藏期间,处理组与对照组草莓果实其可溶性固形物含量也呈先上升后下降的趋势(图1C)。在贮藏期的2~4d,经由1%浓度CaCl2处理的果实其可溶性固形物含量与对照组无显著差异,而经由2%、5%、10%浓度CaCl2处理的果实其可溶性固形物含量均显著地高于对照组(p<0.05)。
2.2 CaCl2处理对灰葡萄孢的控制效果
如表1所示,CaCl2对灰葡萄孢的菌丝生长、孢子萌发以及由其侵染所引发的草莓果实病害均有显著影响,且与CaCl2浓度呈显著负相关关系。其中,CaCl2浓度与菌落直径和孢子萌发率呈线性相关,其相关系数分别为-0.997(p<0.01)和-0.943(p<0.05)。而CaCl2浓度与草莓果实发病率则符合生长曲线模型,其回归模型相关系数为0.956(p=0.011)。

表1 氯化钙处理对灰葡萄孢的控制效果Table 1 The inhibitory effect of CaCl2to B.cinerea
注:同一列内不同小写字母表示处理间差异达到显著水平(p<0.05)。
2.3 CaCl2处理对草莓果实抗病相关物质的影响
2.3.1 木质素含量的变化 图2A所示为CaCl2处理对草莓果实木质素含量变化的影响。由图可知贮藏期间,各个处理组与对照组草莓果实木质素含量变化趋势相同,但高浓度CaCl2处理(5%和10%)可显著提高果实木质素含量(p<0.05),如在贮期第一天,5%和10%CaCl2处理组果实其木质素含量分别为对照组的2.7倍和3.2倍。说明草莓采后经CaCl2处理,能够诱导其在贮藏期间木质素含量的上升。
2.3.2 抗病性相关酶活性的变化 PAL是多酚类抗病物质合成的限速酶,也是苯丙烷代谢途径的关键酶。由图2B可知,在贮期的第一天,四个CaCl2处理组和对照组果实均出现了PAL活性高峰,但高浓度CaCl2处理组(5%和10%)果实其PAL活性均显著地高于其它三组(p<0.05)。在贮藏后期(7~9d),各个CaCl2处理组与对照组果实PAL活性均迅速上
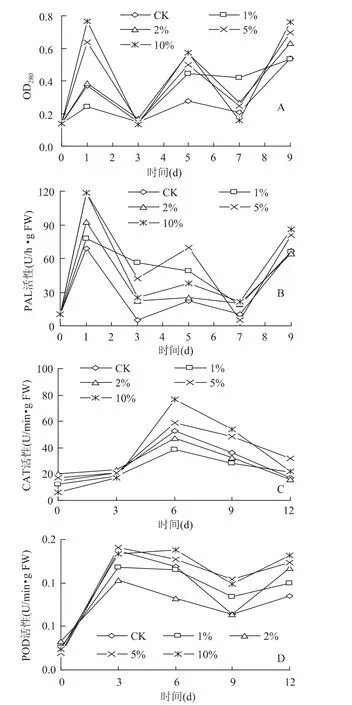
图2 氯化钙浸泡处理对草莓果实(A)木质素含量及(B)PAL、(C)CAT和(D)POD活性的影响Fig.2 Influence of CaCl2soaking on the(A)lignin content,
(B)PAL,(C)CAT and(D)POD activities of strawberry fruit升,这可能是由于贮藏后期病斑直径扩展所致,但各组之间并无显著差异。
CAT和POD是植物体内重要的抗病性相关酶。如图2C所示,贮藏期间,草莓果实CAT活性均呈上升趋势,至第6d达到峰值后迅速下降。高浓度CaCl2处理极大的诱导了CAT活性的上升,在贮期的第6d,10%CaCl2处理组果实其CAT活性为对照组的1.44倍。由图2D可知,各组草莓果实POD活性在贮藏前期(前3d)均呈上升趋势,之后缓慢下降,贮藏后期略有上升,同样可能是由于果实病斑直径扩展所致。在整个贮藏期间,高浓度CaCl2处理组(5%和10%)果实其POD活性始终保持在较高水平,贮藏后期(9~12d)更是显著地高于对照(p<0.05)。
3 讨论
本研究表明草莓果实在贮藏期间品质不断下降,而CaCl2处理能够显著延缓品质下降速度,这与前人的研究结果相一致[5-6]。经10%和5%浓度的CaCl2处理后,草莓果实维生素C下降幅度显著小于对照(图1A),可滴定酸和可溶性固形物含量在贮藏后期(2~4d)也均显著的高于对照(图1B、图1C),这主要与钙离子能够稳定细胞壁以及细胞膜有关[7]。此外,CaCl2处理对灰霉菌的生长有明显抑制作用,能显著降低灰葡萄孢菌的菌落直径和孢子萌发率,而在体内试验中,经CaCl2处理过的草莓其灰霉病发病率显著低于对照组(表1)。这说明CaCl2处理能够减缓采后草莓品质下降速度并抑制灰霉病的发生。
苯丙烷代谢途径是植物体产生木质素和植保素等抗病害物质的重要途径,PAL经解氨作用将莽草酸途径产生的L-苯丙氨酸生成反式肉桂酸从而进入苯丙烷代谢。之前研究结果表明,当植物受到诱导和伤害时防卫系统被激活,PAL活性会迅速上升,同时产生大量抗病性物质如植保素和木质素[8]。而木质素的合成被认为是植物体内主动防御的表现形式,同时木质素可以通过木质化细胞壁组织达到抗病菌的目的,是抗病物质之一。Ca2+是诱导主动防御的激发子之一,此外,Ca2+被认为是植物细胞内调节代谢的第二信使,能参与植物体与病害之间的相互作用[9]。本研究结果表明,在较高浓度CaCl2(10%和5%)影响下,采后草莓PAL活性迅速上升,与对照组相比差异显著(图2B),同时果实木质素含量(图2A)与抗病害能力(表1)显著上升,确实起到了防御病害的目的。
CAT和POD是重要的抗病性相关酶,能够调节植物果实的活性氧代谢[10]。活性氧爆发是果实被病原菌侵染后所产生的早期反应之一,其在植物体内有重要的抗病作用,可以直接杀死病原菌,但过高的活性氧浓度同时会伤害植物自身细胞,因此,诱导产生的活性氧应尽快清除。而CAT可以酶解H2O2,从而调节活性氧浓度,POD也是清除H2O2的主要酶类之一[11-12]。之前有研究表明,Ca2+是活性氧爆发的信号分子,植物体受到侵害时活性氧爆发,同时引起特异性钙波,激发植物体防卫功能[13]。本研究结果表明,在贮藏中后期(6~12d),10%和5%浓度的CaCl2处理组草莓果实其CAT和POD活性均不同程度的增加(图2C、图2D),这会增强果实对于活性氧的清除能力,因而可以避免CaCl2诱导所产生的活性氧同时伤害果实自身细胞,有助于延缓果实老化、保持果实品质。
4 结论
本试验将草莓果实在10℃条件下贮藏4d,研究发现CaCl2处理能够有效延缓果实维生素C、可滴定酸及可溶性固形物含量下降,保持果实品质;且CaCl2浓度不同,对于草莓果实的保鲜效果不同。综合上述三个品质指标分析,经10%浓度CaCl2处理的草莓果实在贮藏期间品质最好。
CaCl2处理对草莓采后灰霉病有较为明显地抑制作用,且该抑病效果与CaCl2浓度呈正相关。此外,CaCl2处理可以诱导采后草莓抗病相关物质的积累,同样,CaCl2浓度不同,对于果实抗病性的诱导能力不同。综合分析,经10%浓度CaCl2处理的草莓果实在贮藏期间木质素积累最多、抗病性相关酶活性变化最显著。
[1]Maor R,Shirasu K.The arms race continues:battle strategies between plants and fungal pathogens[J].Current Opinion in Microbiology,2005,8:399-404.
[2]Manganaris G,Vasilakakis M,Diamantidis G.The effect of postharvest calcium application on tissue calcium concentration,quality attributes,incidence of flesh browning and cell wall physicochemical aspects of peach fruits[J].Food Chemistry,2007,100:1385-1392.
[3]RodriguesJ,FaixO,PereiraH.Improvementofthe acetylbromide method for lignin determination within large scale screening programmes[J].European Journal of Wood and Wood Products,1999,57:341-345.
[4]曹建康,姜微波,赵玉梅.果蔬采后生理生化实验指导[M].北京:中国轻工业出版社,2007:101-111.
[5]杨福臣.钙处理对草莓保鲜效果的影响[J].贵州农业科学,2010,38(10):180-182.
[6]周绪宝,习佳林,郝建强,等.采前钙处理对采后草莓贮藏品质的影响[J].北京农学院学报,2012,27:18-20.
[7]张秀梅,杜丽清,王有年,等.钙处理对果实采后生理病害及衰老的影响[J].河北果树,2005,1:3-4.
[8]崔建东,李艳,牟德华.苯丙氨酸解氨酶(PAL)的研究进展[J].食品工业科技,2008,30:306-308.
[9]张蓓,刘刚,王冬梅.植物抗病防卫反应中的特异性钙信号[J].细胞生物学杂志,2008,30:611-616.
[10]景红娟,周广舟,谭晓荣,等.活性氧对植物自噬调控的研究进展[J].植物学报,2012,47:534-542.
[11]陈金峰,王宫南,程素满.过氧化氢酶在植物胁迫响应中的功能研究进展[J].西北植物学报,2008,28:188-193.
[12]丁薪源,曹建康.果蔬过氧化物酶酶学特性研究进展[J].食品科技,2012,10:62-66.
[13]Chandra S,Low P S.Measurement of Ca2+fluxes during elicitation of the oxidative burst in aequorin-transformed tobacco cells[J].Journal of Biological Chemistry,1997,272:28274-28280.
猜你喜欢
农业科技通讯(2023年1期)2023-02-12
热带作物学报(2022年7期)2022-08-06
今日农业(2022年4期)2022-06-01
作物学报(2022年4期)2022-02-10
浙江农业学报(2017年1期)2017-05-17
现代检验医学杂志(2016年1期)2016-11-12
新农业(2016年20期)2016-08-16
中国蔬菜(2015年9期)2015-12-21
山东医药(2015年38期)2015-12-07
医学研究杂志(2015年4期)2015-06-10
