
长白山次生杨桦林林分结构
2013-08-29 09:29夏富才赵秀海彭道黎倪瑞强汪金松
华南农业大学学报 2013年3期
夏富才 ,赵秀海 ,彭道黎 ,倪瑞强 ,汪金松,范 娟
(1 北华大学 林学院,吉林 吉林 132013;2 北京林业大学 森林资源与生态系统过程北京市重点实验室,北京 100083)
林分空间结构是指树木在林地的分布格局及其属性在空间上的排列方式,表达的是树木之间树种、分布、大小等空间关系[1].林分空间结构的研究对深入了解森林植被与环境的关系,以及森林的生长、发育、更新及演替规律具有重要意义.在经典森林经理学的研究方法中,通常以树种多样性、年龄、树高、直径、形数、林层、密度和蓄积等一维参数来指示林分的结构,很难满足林分动态监测以及森林近自然经营的需要[2].在传统林分结构分析方法的基础上,惠刚盈等[3-6]提出了以相邻木为单元来构建林分空间结构参数,能更好地表达林分在水平方向和垂直方向的异质性.3 个空间结构参数(混交度、大小比数和角尺度)分别描述树种组成及空间配置情况、树种生长优势度和树木水平分布格局.以相邻木关系为基础的方法充分考虑了点的空间位置,能获取种群数量分布的空间信息,分析林木在空间结构单元中的空间关系,林分空间结构的可解析性较高.
近百年来对长白山森林的不合理采伐使原始林遭到严重损害,在当地形成了许多质量低下的次生林.如何保护和恢复当地天然次生林已成为亟待解决的问题.准确获取可解析性强的次生林空间信息是实现长白山森林近自然经营、可持续发展的基础.到目前为止,有少数文献应用与林木有关的结构多样性指数分析了长白山云冷杉针阔混交林和天然红松阔叶林的空间结构[7-9],可至今还缺乏对采伐干扰形成的次生杨桦林的林分结构研究.本研究参考热带雨林生物多样性监测规范,在长白山北坡建立了面积达5.2 hm2的次生杨桦林样地,对样地内的每株林木进行定位监测,用相邻木结构单元法分析了该林分的空间结构,以此为当地次生林的可持续经营提供科学参考.
1 试验地概况
本研究地设在长白山白河林业局光明林场,林分在20 世纪50年代前遭受过零星的盗伐,在1976年经历过一次强度为18%的轻度择伐.林分立地平缓,排水良好.研究区属于温带大陆性山地气候,受东太平洋季风的影响,夏季温暖,雨量充沛,冬季酷寒,平均气温3.3℃,8月最热,平均气温20.5℃,1月最冷,平均气温为-16.5℃.年均降水量600~900 mm,主要集中在6—8月.土壤为发育在火山灰上的深棕色森林土.海拔748 m.地理坐标为N42°20'907″,E128°07'988″.
2 研究方法
2.1 样地调查及林分基本情况
2005年7月,在吉林省白河林业局光明林场次生杨桦林所在地,用TOPCON 全站仪建立了1 个200 m×260 m 的永久样地,对样地内胸径(DBH)大于1 cm 的林木个体测量树高及横、纵坐标,以此确定林木的空间位置.样地共有树种34 种,林分密度为3 157 株/hm2,平均胸径为11.8 cm,树种组成为3 紫椴Tilia amurensis+2 白桦Betula platyphylla +1 红松Pinus koraiensis+1 臭冷杉Abies nephrolepis +1 水曲柳Fraxinus mandshurica+1 青杨Populus ussuriensis+1 蒙古栎Quercus mongolica +色木槭Acer mono +枫桦Betula costata+响杨Populus davidiana+长白鱼鳞松Picea jezoensis +紫花槭Acer pseudo-sieboldianum +山槐Maackia amurensis.
2.2 分析方法
应用空间结构分析法,从3 个方面解析林分空间结构:1)林分的非同质性,即林木组成和空间配置情况;2)林木的非均一性,即林木的生长优劣程度;3)林木的非规则性,即林木个体在水平方向上的分布格局.本文用大小比数描述林木大小分化程度,用混交度描述林木空间配置,并用角尺度描述林分水平格局[10].
2.2.1 树种混交度 树种混交度(M)定义为与目标树i 最近的4 株相邻木中与目标树不同种个体所占的比例,Mi为某一株目标树的混交度,用公式表示为:
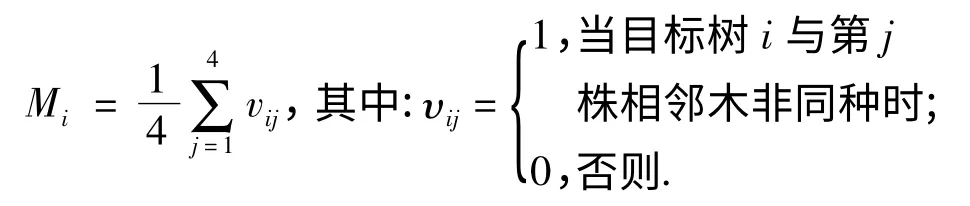
Mi=0 时,表示零度混交;Mi=0.25 时,表示轻度混交;Mi=0.50 时,表示中度混交;Mi=0.75 时,表示强度混交,Mi=1.00 时,表示完全混交.
2.2.2 大小比数 大小比数(U)是指胸径、树高或冠幅大于目标树的相邻木占最近4 株相邻木的株数比例,Ui为某一株目标树的大小比数,用公式表示为:
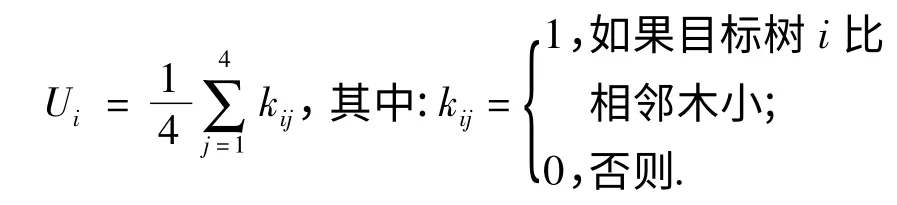
Ui=0 时,表示目标树处于绝对优势地位;Ui=0.25 时,表示目标树处于优势地位;Ui=0.50 时,表示目标树处于中庸地位;Ui=0.75 时,表示目标树处于劣态;Ui=1.00 时,表示目标树处于完全劣态.
2.2.3 角尺度 角尺度(W)可用来描述相邻木围绕目标树的均匀程度.任意2 个邻接最近相邻木的夹角有2 个,小角为α,最近相邻木均匀分布时的夹角设为标准角α0.则角尺度被定义为α 角小于标准角α0的个数占所考察的4 个夹角的比例.Wi为某1株目标树与最近4 株相邻木构成的角尺度值,表达式为:

3 结果与分析
3.1 林木种间关系
据调查,次生杨桦林样地共有34 个树种.运用林分空间结构分析软件winkelmass 得到该林分平均混交度为0.63,其中完全混交、强度混交、中度混交、轻度混交和零度混交的占比分别为34%、24%、17%、12%和13%,说明该林分整体上处于中度以上混交水平.又因林木的株数、大小、繁殖及更新等不同,树种间的混交度特点差别很大(表1).位于林分下层,以营养繁殖为主的林木混交度偏低.如,簇毛槭弱度混交和零度混交的株数占比合计为62%,说明该树种空间隔离程度低,成簇现象明显.紫花槭在该林分中株数最多,占整个群落的22%,各种混交度的占比接近,它的空间隔离程度也很低.青楷槭Acer tegmentosum 和花楷槭A.ukurunduense 整体上处于中度混交水平,各种隔离程度的林木个体相对均一.位于林分中、上层树种的平均混交度都在中度混交以上,混交度水平普遍高于下层树种.水曲柳是林冠上层林木中混交度水平最低的树种,弱度混交和零度混交的株数占比之和将近30%,这可能与该树种更新良好和存在大量幼年个体有关.在该林分的针叶树中,臭冷杉的个体数最多,占林分总株数的17%,且幼苗和小树较常见,约20%的个体处于弱度混交及以下水平,强度混交及其以上混交水平的比例占60%.其余的乔木,除糠椴Tilia mandshurica 外,绝大多数个体的低强度混交水平的情形很少见,中度混交水平的比例均小于3%,因此,这些树种的平均混交度水平都很高.林分中能达到完全混交度水平的树种均是株数很少的稀见树种,如山梨Pyrus ussuriensis、响杨等.

表1 次生杨桦林树种混交度及其取值分布Tab.1 Species' mingling degree and their distribution in secondary popular-birch forest
3.2 林木大小分化


表2 次生杨桦林树种大小比数及其取值分布Tab.2 Neighborhood comparison and its distribution of tree species in secondary popular-birch forest
3.3 林木水平分布格局


图1 次生杨桦林林分角尺度分布Fig.1 The value distribution of neighborhood pattern in secondary popular-birch forest
4 讨论
长白山次生杨桦林林分空间结构特征与采伐干扰造成的次生演替以及各树种在林分中的生长状况、繁殖方式、更新策略等有关.
林分的整体混交度水平与群落的干扰历史及所处的演替阶段等有关[11].次生杨桦林的混交度水平明显高于当地的红松阔叶林[12]以及遭受强度采伐干扰后形成的次生白桦林[13],这很可能是遭受采伐干扰的林分中,大径木被伐除,相同物种的最近相邻个体得到抚育,林分的混交度水平被人为提高,又由于先锋树种的侵入和定植增加了该群落的乔木树种丰富度,进一步提高了林分的混交度水平.因此,并非所有的次生林的混交度水平都低于原始林,混交度水平高的森林也不一定是材积高、稳定性高的林分.在调整次生杨桦林以及次生白桦林林分混交度时,应适当减少先锋树种的比例,同时充分考虑其他树种生长、繁殖特性及参考当地老龄林的混交度特点.
与当地原始红松阔叶林相比,次生杨桦林中生长上占优的树种以先锋树种为主,而有可能达到林冠上层的红松、紫椴、水曲柳等在生长上并不完全处于优势地位,而是分布在林分的各个层次.这说明次生杨桦林还处于演替的早期阶段,林冠中层个体之间竞争激烈.因此,在林分改造中,参考原始林树种构成的特点,适当提高当地顶级群落优势树种的优势度,防止次生林进一步退化,从而促进次生林的正向演替.
在原始森林中,由于自疏作用和种内、种间竞争及生态位分化,使林木之间更合乎随机化分布趋势[14].而次生杨桦林由于受到人为采伐干扰,林木原有格局已被改变,加之一些先锋树种如白桦、青杨的侵入,小径级林木群团状特点突出,导致林分在总体上呈现聚集分布的格局,这与当地相似类型的次生白桦林的分布格局一致[13].
在今后针对该类次生林的经营管理中,建议以优化和构建合理的林分空间结构为出发点,调整林分的角尺度,将角尺度偏小的林木作为调整对象,减少林分中林木团状分布的比例,促进林分向天然林林分随机分布趋势发展.在经营中保留有助于调节混交度的未来顶级群落的针、阔叶优势树种,使次生林林分混交度水平逐渐接近原始林.有意识地调整林木个体的大小分化水平,通过抚育间伐、人工促进、天然更新等措施,为珍贵幼树提供生长空间,逐步降低林分中非目的树种的中、小径木的比例,使林分径级分布趋于合理[15].通过以上经营管理措施改善树种组成以及竞争关系,调整林分的分布格局,促进进展演替,诱导次生林向原始林的结构特征发展.
[1]惠刚盈,胡艳波,赵中华.再论“结构化森林经营”[J].世界林业研究,2009,22(1):14-19.
[2]MACKINNON A,TROFYMOW J A.Structure,processes,and diversity in successional forests of coastal British Columbia[J].Northwest Sci,1998,72(S2):1-3.
[3]惠刚盈.角尺度:一个描述林木个体分布格局的结构参数[J].林业科学,1999,35(1):39-44.
[4]惠刚盈,von GADOW K,ALBERT M.一个新的林分空间结构参数:大小比数[J].林业科学研究,1999,12(1):4-9.
[5]惠刚盈,胡艳波.混交林树种空间隔离程度表达方式的研究[J].林业科学研究,2001,14(1):23-27.
[6]惠刚盈,李丽,赵中华,等.林木空间分布格局分析方法[J].生态学报,2007,27(11):4717-4728.
[7]胡艳波,惠刚盈,戚继忠,等.吉林蛟河天然红松阔叶林的空间结构分析[J].林业科学研究,2003,16(5):523-530.
[8]安慧君,惠刚盈,郑小贤,等.不同发育阶段阔叶红松林空间结构的初步研究[J].内蒙古大学学报:自然科学版,2005,36(6):116-120.
[9]贾秀红,郑小贤.长白山过伐林区云冷杉针阔混交林空间结构分析[J].华中农业大学学报,2006,25(4):436-440.
[10]徐海,惠刚盈,胡艳波,等.天然红松阔叶林不同径阶林木的空间分布特征分析[J].林业科学研究,2006,19(6):687-691.
[11]WELLS M L,GETIS A.The spatial characteristics of stand structure in Pinus torreyana[J].Plant Ecology,1999,143(2):153-170.
[12]夏富才,赵秀海,潘春芳,等.长白山阔叶红松林林分空间结构[J].应用与环境生物学报,2010,16(4):529-533.
[13]夏富才,赵秀海,潘春芳,等.长白山白桦林空间结构研究[J].西北植物学报,2011,31(2):407-412.
[14]王蕾,张春雨,赵秀海.长白山阔叶红松林的空间分布格局[J].林业科学,2009,45(5):54-59.
[15]赵中华,袁士云,惠刚盈,等.甘肃小陇山5 种不同灌木林改造模式对比分析[J].林业科学研究,2008,21(2):262-267.
猜你喜欢
阿来研究(2020年1期)2020-10-28
现代农业研究(2017年11期)2018-01-12
辽宁林业科技(2017年4期)2017-06-22
辽宁林业科技(2017年4期)2017-06-22
现代园艺(2017年20期)2017-02-03
新世纪水泥导报(2016年1期)2016-07-01
中央社会主义学院学报(2016年2期)2016-05-04
土木建筑工程信息技术(2013年1期)2013-10-17
