
急性胎兔宫内窘迫模型脑组织病理及NMDAr1表达实验研究
2013-08-21 07:44林也容林进皇吴芳芳郁毅刚
中国医药科学 2013年23期
林也容 林进皇 吴芳芳 郁毅刚▲
1.解放军第一七五医院暨厦门大学附属东南医院妇产科,福建漳州 363000;2.解放军第一七五医院暨厦门大学附属东南医院神经外科,福建漳州 363000
胎儿在宫内有缺氧征象危及胎儿健康和生命者,称为胎儿窘迫(fetal distress,FD)[1],是胎儿围产期死亡及新生儿神经系统后遗症的常见原因,占围产儿死亡原因的首位[2],给社会、家庭和个人带来沉重负担。深入研究FD的发病机制,包括病理生理机制和分子机制,具有重要的意义。本研究通过建立急性胎兔宫内窘迫模型,观察研究其胎兔脑组织病理及NMDAr1表达情况。
1 材料与方法
1.1 动物及分组
健康成年新西兰大白兔4周左右孕兔,普通级。体质量3.2~3.7kg,由上海松江松联实验动物场生产,厦门大学医学院动物中心提供。动物许可证:SCXK(沪)2007-0011。实验动物使用单位许可证编号:SYXK(军)2002-47。动物实验人员资质许可证编号:军动管字第2006E01028。实验孕兔饲养3d,每日傍晚测体温、脉搏、呼吸1次。至孕32~34d实验,剔除异常极值动物,经统计分析,个体间无明显差异。编号按随机表分成3组,缺血模型组9只,假手术组9只,空白组12只。
1.2 实验方法
模型制作:妊娠34d孕兔以2.5%戊巴比妥钠0.1mL/100g进行腹腔麻醉后,耻区下腹部正中切口打开腹腔,暴露双侧子宫角,检查胎盘、子宫及孕兔活力。分离出进入每个胎盘的分支动脉血管,以无创动脉夹钳夹20min,然后松开血管夹,恢复血流,建立急性胎兔宫内窘迫模型。缺血模型组孕兔行开腹子宫动脉结扎术,建立胎兔宫内窘迫缺血缺氧模型;假手术组孕兔行开腹术,不结扎子宫动脉;空白组孕兔不行任何手术处理。除空白组胎兔直接断头取脑外,其他组胎兔均在手术处理后6h断头处死、取脑,固定,显微镜下观察各组胎兔脑组织病理形态及NMDAr1表达情况。
1.3 脑组织病理检测
取4%甲醛固定脑组织,石蜡包埋,切片行HE染色,光镜下观察形态学变化,由同一病理医生采用盲法评价。观察脑组织结构层次是否完整,神经细胞肿胀、变性、坏死,间质出血、水肿等情况。
1.4 脑组织NMDAr1免疫组化表达的测定
取4%甲醛固定脑组织,石蜡包埋,连续切片(4μm),每例标本切5张免疫组化染色用。采用改进型SP法进行免疫组化染色,严格参照试剂盒说明操作。试剂盒型号,购于XX公司。每次染色时均用已知阳性切片作阳性对照,用PBS(磷酸缓冲液,0.01mol/L,pH值7.2)代替一抗作阴性对照。NMDAr1表达均位于胞质和胞核内,表现为浅黄、棕黄或棕褐色。用半定量法评估其染色,无阳性细胞数为(-),阳性细胞数 25% 以下为(+),25%~50% 为(++),51%~75%为(+++),超过 75% 为(++++)。
2 结果
2.1 脑组织HE染色结果
HE染色空白组胎兔脑组织切片示,脑皮质各层发育正常,其中分子层,内颗粒层、外颗粒层以及锥体细胞层均匀致密,完整;锥体细胞数量中等,胞体较小致密,轴突干净完整、延伸,胞核居中。白质间少量血管,肌浆红染,胶质丰富;血管腔内可见红细胞(图1~2)。假手术组脑组织切片HE显示,各层结构存在,锥体细胞数量中等,基本同空白组。胞体稍肿胀,未见明显空泡变性,轴突完整,光滑延伸;细胞间偶见少量红细胞,无明显间质炎细胞浸润(图3~4)。HE染色缺血模型组切片示,胎兔脑皮层组织结构层紊乱,部分坏死崩解,分界不清;锥体胞胞体肿胀,数量稀少,单细胞体积明显增大,形态不规则,边缘较毛糙,胞膜胞质融合。轴突断裂崩解,白质稀疏;间质充血水肿,散在红细胞及浆细胞浸润(图 5~6)。

图1 空白组HE染色(×100)
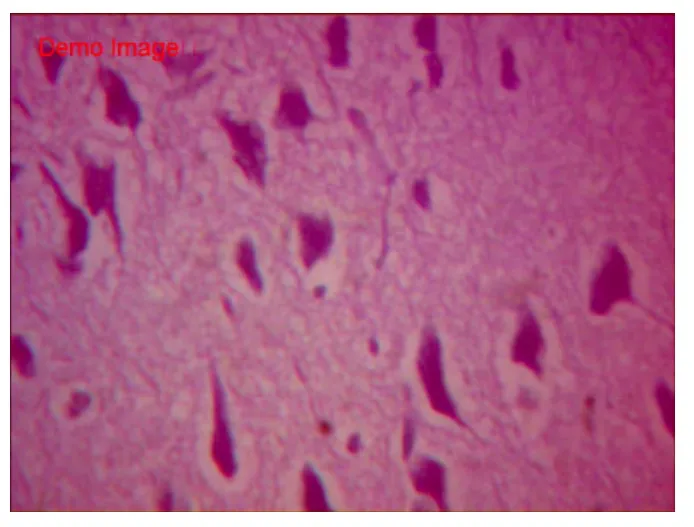
图2 空白组HE染色(×400 )
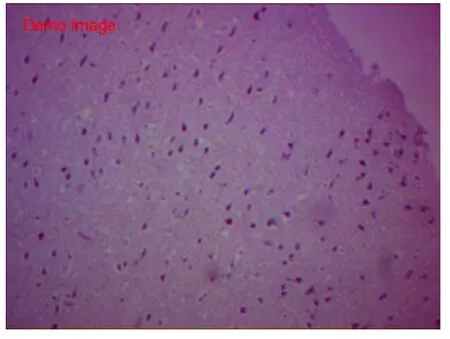
图3 假手术组HE染色(×100)
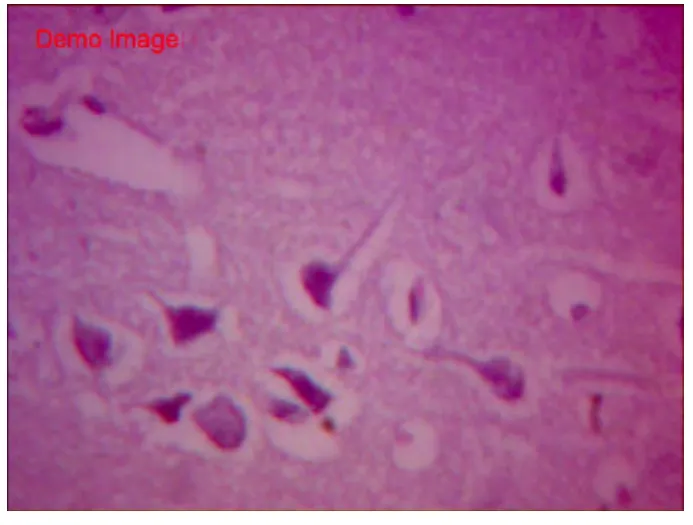
图4 假手术组HE染色(×400)

图5 缺血模型组HE染色(×100)
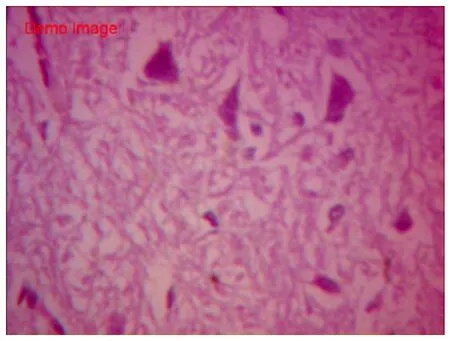
图6 缺血模型组HE染色(×400 )
2.2 脑组织NMDAr1免疫组化的表达
各组胎兔脑切片神经元锥体细胞均有NMDAr1表达,主要在神经元膜表面表达,免疫反应强度有差异。空白组见神经元形态完整、胞膜淡染,NMDAr1蛋白少量表达(+)(图7~8)。假手术组,与空白组基本一致,胞体结构完整,数量中等,细胞膜不典型黄染(+),NMDAr1少量表达(图9~10)。缺血模型组可见神经元坏死崩解,少量残存神经元形态粗糙,胞体肿胀,体积巨大,胞核溶解,大部分胞质、胞膜均黄染,色深;NMDAr1大量表达(+++)(图11~12)。空白组和假手术组NMDAr1免疫反应强度均低于缺血模型组,缺血模型组NMDAr1蛋白呈神经元膜及胞浆内强性表达 。

图7 空白组Nr1组化(×100)

图8 空白组Nr1组化(×400)
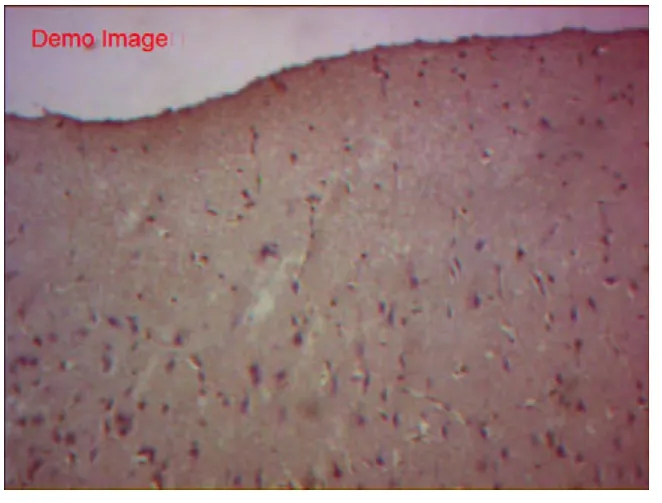
图9 假手术组Nr1组化(×100)

图10 假手术组Nr1组化(×400)

图11 缺血模型组Nr1组化(×100 )

图12 缺血模型组Nr1组化(×400)
3 讨论
胎儿宫内窘迫是由于胎儿缺血缺氧所引起,可发生在临产后,也可发生在妊娠期,是导致死胎死产主要原因之一[3],可分为急性胎儿窘迫和慢性胎儿窘迫[4]。研究急性宫内窘迫脑组织病理形态有助于脑缺血缺氧性损伤病理生理机制的深入了解,对该病的积极治疗和改善预后等发展亦具有相当重要的意义。
本实验研究中,我们选择了与人解剖及生理相似程度较高的兔子作为实验对象,通过对缺血模型组孕兔行开腹子宫动脉结扎术,建立急性胎兔宫内窘迫模型。在术后6h断头取脑,固定后HE染色,显微镜下发现胎兔脑皮层组织结构层紊乱,部分坏死崩解,分界不清;锥体胞体肿胀,数量稀少,单细胞体积明显增大,形态不规则,边缘较毛糙,胞膜胞质融合。轴突断裂崩解,白质稀疏;间质充血水肿,散在红细胞及浆细胞浸润。而只行开腹术不行子宫动脉结扎的假手术组胎兔脑病理组织形态与缺血模型组差异明显,而与空白组基本相同,脑皮质各层结果存在,其中分子层,内颗粒层、外颗粒层以及锥体细胞层均匀致密,完整;锥体细胞数量中等,胞体稍肿胀,未见明显空泡变性,轴突完整,光滑延伸;细胞间偶见少量红细胞,无明显间质炎细胞浸润。本实验中,缺血模型组胎兔的神经病理改变情况与郭世杰等[5]研究报告的相似,所以该模型可成功复制急性宫内窘迫症,病理形态表现明显,能够帮助我们更好观察研究其病理生理过程。
除了病理观察,我们还对胎兔脑神经元锥体细胞进行免疫组化染色,发现各组均有NMDAr1表达,主要在神经元膜表面表达,免疫反应强度差异较明显。假手术组与空白组基本一致,NMDAr1均少量表达,而缺血模型组NMDAr1大量表达(+++)。缺血模型组NMDAr1免疫反应强度高于空白组和假手术组,其神经元膜及胞浆内呈强阳性。由此不难看出,缺血模型组的病理学损伤伴随着NMDAr1蛋白表达,我们可以推断NMDA受体的高表达和激活导致脑组织缺血缺氧性损伤,这与早期实验研究结果基本一致[6-7]。
NMDA受体由 3种亚基组成: NR1、NR2(A-D)和NR3(A-B)[8]。NR1是能单独构成离子通道的功能亚基,而NR2和NR3是不能单独构成离子通道的调节亚基,分别增强或抑制NR1亚基的功能。NR1和NR2的表达是NMDA受体发挥功能所必需的,NR3则不是NMDA受体发挥功能所必需的[9]。因此,NMDArl蛋白表达的变化可反映NMDAr蛋白表达的变化。胚胎期或新生期脑组织内NMDA受体表达量较其他时期高,这对于脑发育如学习和记忆等非常重要。但当损伤因素存在时NMDA受体的过高表达又可损伤神经系统,因此NMDAR表达具有“双刃剑”作用[10]。本研究也证实了当出现急性宫内窘迫时,胎兔的脑组织内代表NMDA受体的NMDAr1表达显著增高,这是胎兔脑组织损伤的关键机制之一。
急性宫内窘迫常致缺血性脑损伤,其机制错综复杂,NMDA受体的过度激活是其中关键点之一,是损伤后诸多继发性病理改变的初始“钥匙”环节[11]。作为一种兴奋性氨基酸的特异性离子型受体,当NMDA受体被过度激活时,大量Ca2+流入细胞内,进而触发一系列生化过程[12]: 激活依赖Ca2+的蛋白水解酶;激活磷酸脂酶C、A1、A2,使膜磷脂水解,花生四烯酸释放增加;使神经递质DA释放增加;自由基大量形成,线粒体氧化磷酸化脱偶联,细胞呼吸抑制;激活核酸内切酶,引起DNA分解。这些生化过程最终促使了神经元坏死和凋亡等病理改变[13],与本实验的病理结果一致。
综上,通过建立急性宫内窘迫模型,观察到了脑组织NMDAr1表达及病理损伤情况,为NMDA受体过度激活致脑组织损伤提供理论依据,对于进一步深入研究急性宫内窘迫脑组织损伤机制具有重要意义。
[1]彭敏丹.胎儿宫内慢性缺氧69例临床观察[J].临床医学,2009,29 (10):95-96.
[2]陈光英,魏守红,陈英,等.产程中胎儿宫内窘迫636例临床分析[J].中国妇产科临床杂志,2004,5(4):290-291.
[3]柴秀仙.围产儿死亡原因分析及围产保健[J].中国妇幼保健,2011,26(2):206-208.
[4]于敏.胎儿窘迫的临床护理[J].中外健康文摘,2011,8(11):371-372.
[5]郭世杰,程门雪,李玉梅,等.新生儿缺氧缺血性脑病临床与神经病理研究[J].中风与神经疾病杂志,2005,22(5):424-426.
[6]Biondi O,Branchu J,Sanchez G,et al.In vivo NMDA receptor activation accelerates motor unit maturation,protects spinal motor neurons,and enhances SMN2 gene expression in severe spinal muscular atrophy mice[J].J Neurosci,2010,30:11288-11299.
[7]王嘉炜,王汉东.代谢性谷氨酸受体5在神经胶质细胞中的研究进展[J].中华神经外科疾病研究杂志,2013,12(5):470-471.
[8]王公明,田玉科,戴体俊,等. NMDA受体2B亚基:一个潜在的镇痛治疗靶点[J].国际麻醉学与复苏杂志,2006,27(5):309-312.
[9]Paoletti P,Netton J.NMDA Receptor subunits:function and pharmacology [J].Curr Opin Pharmacol,2007, 7(1):39-47.
[10]贺艳,赵峰,马晶,等.当归对宫内缺氧幼年大鼠海马CA3区神经元与学习能力的影响及机制[J].时珍国医国药,2010,21(3):646-649.
[11]郁毅刚,徐如祥.原代培养神经细胞膜经N-甲基-D-天冬氨酸和MK-801作用后原子力显微镜观察[J].第一军医大学学报,2002,22(11):988-991.
[12]唐敬龙,高维娟.谷氨酸NMDA受体与学习记忆的关系[J].中国老年学杂志,2009,29(18):2408-2410.
[13]陈翔.NMDA受体与脑损伤[J].国外医学(生理、病理科学与临床分册),1996,16(3):179-181.
猜你喜欢
西北民族大学学报(自然科学版)(2022年2期)2022-07-06
纺织科技进展(2021年4期)2021-07-22
兰州理工大学学报(2021年3期)2021-07-05
现代装饰(2018年5期)2018-05-26
中国组织化学与细胞化学杂志(2017年1期)2017-06-15
中成药(2017年6期)2017-06-13
中国市场(2017年5期)2017-03-15
中国生化药物杂志(2015年4期)2015-07-07
医学研究杂志(2015年12期)2015-06-10
弹箭与制导学报(2015年1期)2015-03-11
