
野生条石鲷精巢的年周期发育
2013-08-14 05:50:22彭志兰柳敏海傅荣兵张凤萍罗海忠
海洋通报 2013年6期
彭志兰,柳敏海,傅荣兵,张凤萍,罗海忠
(浙江省舟山市水产研究所 浙江省海洋开发研究院水产养殖技术研究推广中心,浙江 舟山 3160001)
条石鲷(Oplegnathus fasciatus,T.&S.)属鲈形目(Perciformes)、鲈总科(Percoidea)、石鲷科(Oplegnathidae),石鲷属(Oplegnathu)(中国科学院动物研究所等,1962;朱元鼎等,1963),俗称日本鹦鹉鱼。主要分布于我国黄海、东海、日本北海道以南等海域(张春霖等,1995),为近海温暖性中下层名贵经济鱼类(Koh et al,1922)。其体态优美,色泽艳丽、肉质细嫩,营养丰富,具有很高的食用和观赏价值(常抗美等,2005),备受国内外消费者青睐,市场前景广阔。在中国,虽然分布区域较广,全年皆产,但始终没有形成明显的盛渔期(肖志忠等,2008)。因此,近年来许多学者对条石鲷胚胎发育、亲鱼培育及繁育技术等进行了大量研究(全汉锋等,2007;柳学周等,2008),但有关野生条石鲷精巢发育的周年变化研究至今还未见报道。研究野生条石鲷精巢发育的组织学,旨在丰富条石鲷繁殖生理学内容,并为完善条石鲷的人工繁育技术提供科学依据。
1 材料和方法
1.1 材料
试验用的条石鲷为舟山嵊山海捕野生鱼,从2007年1月到12月逐月采样,每次取10尾。体长为 15~42 cm,体重为 41~2 850 g,♀∶♂为 1.2∶1。
1.2 方法
测量每尾条石鲷的全长、体长、体重等生物学指标,然后解剖,观察性腺发育,测量性腺、肝脏重量,计算性腺成熟系数(GSI)=[性腺重/(体质量-内脏重)] ×100、肝重指数(HSI)=[肝脏重/(体质量-内脏重)]×100。分别从精巢前、中、后部各取部分组织用Bouin′s液固定,脱水、透明、石蜡包埋,连续切片厚度为5~7 μm,H-E染色,树胶封片,用OLYMPUS显微镜观察、拍照。精巢分期方法根据精巢外观与精巢内生殖细胞的成熟程度变化来划分。
1.3 数据处理
实验所得数据均表示为平均数±标准差(Mean±SE),采用SPSS11.5统计软件包进行数据处理和检验分析,当P<0.05时差异显著。
2 结果
2.1 GSI、HSI的周年变化
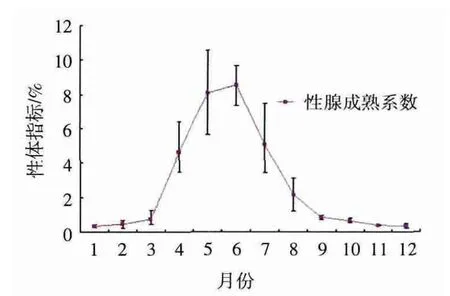
图1 条石鲷雄性GSI的季节变化
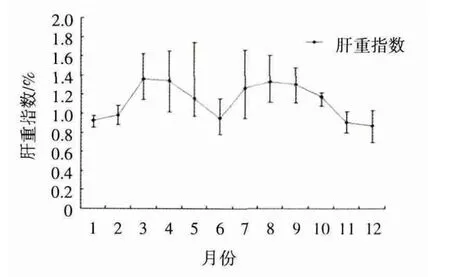
图1 条石鲷雄性HSI的季节变化
野生条石鲷GSI变化如图1。GSI在1-3月是逐渐升高,但各月GSI差异不显著(P>0.05)。4月份GSI急剧升高,与前几个月显著升高(P<0.05)。在 5-6月 GSI达到峰值,分别为 8.11±1.95、8.57±0.90,与其他月份的GSI值均显著高(P<0.05),从7月份开始GSI逐渐下降,7月份的GSI值除与4月份的差异不显著外(P>0.05),与其他月份均差异显著(P<0.05)。GSI在12月份达到最低值,但与9-11月均差异不显著(P>0.05)。
HSI平均值在1-3月是逐渐在上升的,且各月间GSI值差异显著(P<0.05)。从4-6月HSI平均值是逐渐下降。7-8月HIS又开始上升的,从9月开始HIS又逐渐下降,到12月达到最低,为0.87±0.13。
2.2 精巢的形态特征
精巢位于腹腔背面,为一对伸长的器官。精巢外包一层薄的结缔组织,生殖上皮随着结缔组织向内延伸,在精巢内形成许多管状的精小管,这些精小管在精巢内互相连接成网状,最后汇集成一条大的输精管开口在直肠与输尿管之间的泄殖乳突上(图板Ⅰ-1、Ⅰ-2)。条石鲷精巢属于叶状结构(lobular structure),生殖细胞沿着精小管分散分布,精子游离在中央腔,在生精小管内有许多包囊组成。随着精巢的生长发育,在精小管内可见各个包囊生精细胞的发育不是同步的,但在同一个包囊中的生精细胞的发育则是同步的,因此,条石鲷属于分批同步排精类型。
2.3 精巢发育的组织学
2.3.1 第Ⅰ期精巢
该期见早期发育精巢,外观为细线状,边缘光滑。腹膜和肠系膜及腹壁紧密相连。精原细胞分布在生精小管中。从组织切片观察可见,多个精原细胞被结缔组织包裹在一起,呈长柱形或不规则形状,细胞沿管状周排列。精原细胞胞质嗜碱性,着色深,细胞体积大,椭圆形,胞径20.79~24.46 μm,胞核很大,核径 7.39~12.86 μm,核居中或偏于一侧,有一个核仁,着色深,核质分布均匀,核膜清晰,保质着色浅或几乎不着色(图板Ⅰ-3、Ⅰ-4)。Ⅰ期精巢一生只出现一次。
2.3.2 第Ⅱ期精巢
精巢稍微增大,边缘波浪形,灰白色,血管不明显。精巢占腹腔约1/10的体积,位于腹腔后部。此期精巢分繁殖后再待成熟个体和初次进入生殖周期的个体。精小叶蜂窝状界限明显,胞核明显,细胞膜逐渐不清晰,染色仍然很深(图板Ⅰ-5)。初级精母细胞为椭圆形,直径比精原细胞小,为7.24~10.86 μm,染色质丰富,染色比精原细胞深,没有明显的核仁。
2.3.3 第Ⅲ期精巢
精巢扁片状,边缘波浪形,灰白色。精巢占腹腔约2/5的体积,位于腹腔后部和上部。生殖细胞数量进一步增加,Ⅲ期早期生殖细胞正经历由精原细胞到精母细胞的分裂活跃阶段,初级精母细胞个体体积变小,胞径6.50~10.29 μm,核质比增大,胞核染色加深。Ⅲ期晚期由初级精母细胞经历第一次分裂所形成的次级精母细胞体积更小,嗜碱性增强,核染色极深,胞径2.56~4.35 μm。此时的Ⅲ期精巢精巢表现出非同步性更明显,精原细胞、初级精母细胞、次级精母细胞同时存在(图板Ⅰ-6)。
2.3.4 第Ⅳ期精巢
精巢呈乳白色,表面血管清晰(图板Ⅰ-1),精巢占腹腔1/2以上的体积。Ⅳ期精巢发育的同步性增强,特别是晚期,精子细胞和正在形成的精子充满每个精小囊(图板Ⅰ-7),轻压腹部有少量精液流出。精子细胞为圆形小粒状,体积很小,直径为1.71~2.70 μm,细胞核大,染色极深。此期的生精小管开始退化消失。
2.3.5 第Ⅴ期精巢
精巢膨大,较软,乳白色,表面血管更清晰,精巢占腹腔2/3以上的体积。此时细胞数量达到顶峰,整个精巢充满成熟精子,精小囊消失或融合相通,精子密集,被染成浅红色的尾巴可形成流线形,而精子头部被染成蓝色的圆形。此期轻按鱼体腹部大量乳白色精液从泄殖孔流出。
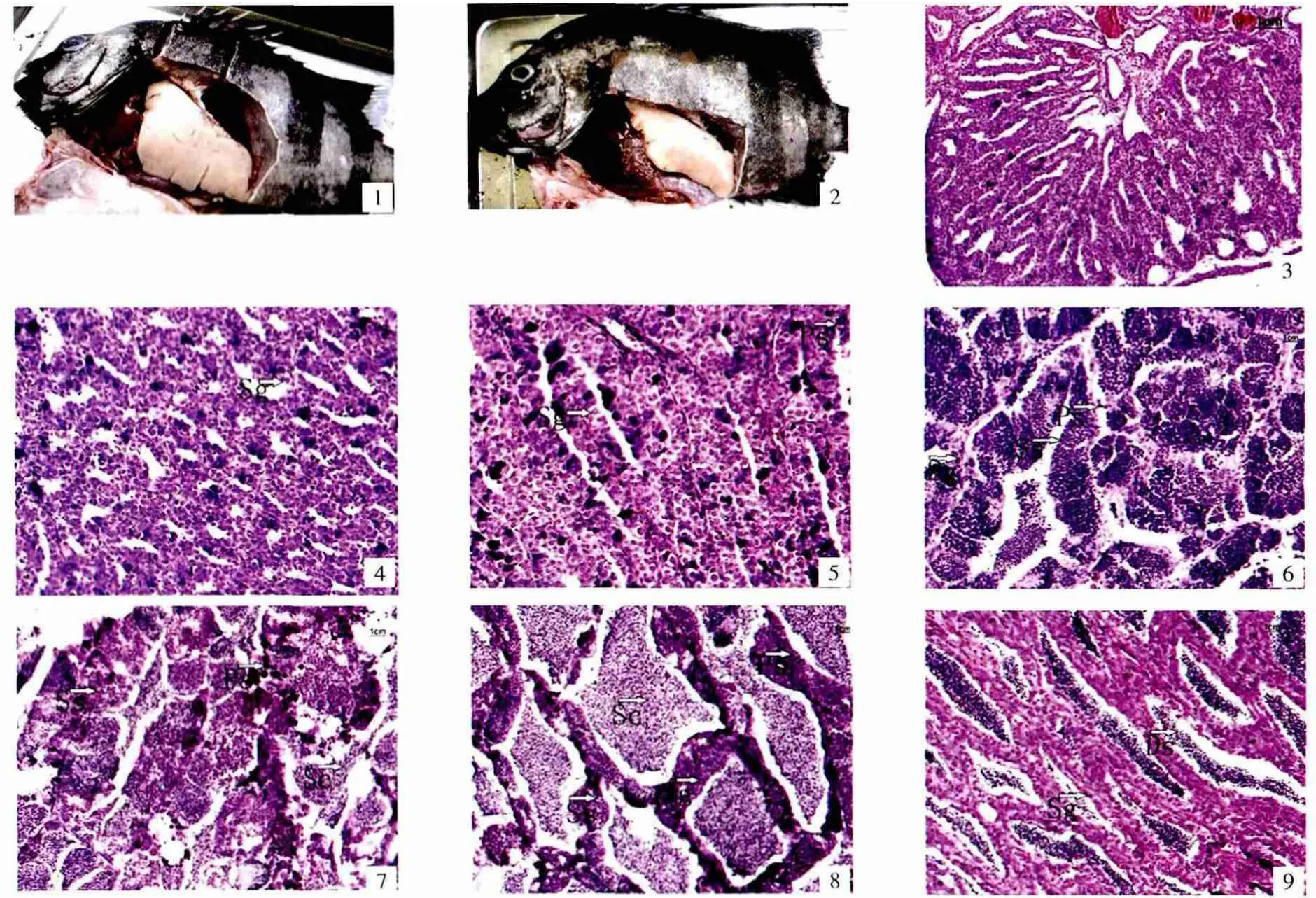
图板Ⅰ 条石鲷精巢年周期发育的组织学观察
2.3.6 第Ⅵ期精巢
经过排出精子后的精巢体积缩小(图板Ⅰ-2),但切片显示,仍然有大量的精子细胞和精子(图板Ⅰ-8),同时精母细胞、精原细胞的数量增加。且这个过程时间较长,从7月份开展持续到9月份。到9月份时,切片显示精囊萎缩,精子排出留下许多囊腔,有些囊腔内残留一些未排出的精子,除了退化的精子细胞外,主要是大量的精原细胞(图板Ⅰ-9)。
3 讨论
条石鲷雄性GSI从4月份开始急速上升,最高值出现在5月份,GSI为10.56%,最高平均值出现在6月份,为8.57±0.90,且5-6月GSI与其他月份的GSI值均显著高(P<0.05)。这与组织学染色发现处于此期的野生条石鲷精巢大多处于Ⅳ~Ⅴ的结果相吻合。GSI从7月开始降低,这也与组织学染色发现处于此期的野生条石鲷精巢大多处于Ⅵ的结果相吻合。
HSI的变化呈波浪型变化,1-3月是逐渐在上升,从4-6月逐渐下降,7-8月HSI是上升的,从9月开始HSI逐渐下降。HSI的变化与性腺的发育是紧密联系的,条石鲷雄鱼性腺开始成熟时HSI缩小,这与鲇(Silurus asotus)(朱玲等,2002)、石鲽(Kareius bicoloratus)(高玲 等,2006)雄鱼性腺开始成熟时HSI缩小的结果相一致,这可能由于雄鱼不存在卵黄蛋白原的合成和转移有关。鲇和石鲽在繁殖期后期HSI还是下降的,而条石鲷在7-8月HSI是上升的,从9月开始HIS才下降,这与鲇和石鲽的变化趋势不同,笔者认为这可能是繁殖季节以及繁殖方式的差异造成的。
野生条石鲷雄鱼为分批同步排精类型,精巢发育与其他硬骨鱼类相似,生殖细胞经历精原细胞、初级精母细胞、次级精母细胞、精子细胞和精子5个典型的发育阶段。条石鲷精细胞在发育过程中是非同步的,特别在Ⅲ期精巢非同步性更明显,精原细胞、初级精母细胞、次级精母细胞同时存在。这与石鲽(高玲等,2006)、鳗鲡(Anguilla japonica)(林鼎等,1984)雄性性腺发育的组织学分析相似。在鳗鲡与石鲽的研究中发现精细胞的发育存在“同步—不同步—同步”的现象,在条石鲷的组织学研究中也同样存在,在Ⅲ期精巢非同步性非常明显,但到了后期,这种非同步性逐渐消失,慢慢的出现同步性,这为条石鲷分批同步排精提供了保障。
Koh J N,Kim Y U,1992.Embryonic development and morphology of larvae and juveniles of parrot fish Oplegnathus fasciatus(Temminek et Schlegel).Bulletin of National Fisheries University of Pusan(Natural Sciences),32(2):29-45.
常抗美,毛建平,吴剑锋,等,2005.条石鲷胚胎及仔稚鱼的发育.上海水产大学学报,14(4):401-405.
高玲,温海深,马学坤,等,2006.野生石鲽精巢发育及注射LHRH-A对血浆T水平的影响研究.中国水产科学,13(5):845-850.
林鼎,林浩然,1984.鳗鲡繁殖生物学研究Ⅲ.鳗鲡性腺发育组织学和细胞学研究.水生生物学集刊,8(2):157-170.
柳学周,徐永江,王妍妍,等,2008.条石鲷的早期生长发育特征.动物学报,54(2):332-341.
全汉锋,肖治中,2007.条石鲷人工繁育技术研究.台湾海峡,26(2):295-300.
肖志忠,郑炯,于道德,等,2008.条石鲷早期发育的形态特征.海洋科学,32(3):25-30.
张春霖,成庆泰,郑葆珊,等,1995.黄渤海鱼类调查报告.北京:科学出版社,153-154.
中国科学院动物研究所,中国科学院海洋研究所,上海水产大学,1962.南海鱼类志.北京:科学出版社,576-577.
朱玲,温海深,毛玉泽,2002.野生鲇性腺发育及其与肝重指数关系的研究.水利渔业,22(5):26-27.
朱元鼎,张春霖,成庆泰,1963.东海鱼类志.北京:科学出版社,254-255.
猜你喜欢
星星·诗歌理论(2023年12期)2024-01-06 08:24:53
中国兽医学报(2022年3期)2022-07-25 06:35:18
数学大王·中高年级(2020年4期)2020-04-19 10:07:16
数理化解题研究(2019年34期)2019-12-19 02:14:46
录井工程(2017年4期)2017-03-16 06:10:37
录井工程(2017年4期)2017-03-16 06:10:36
中国教育技术装备(2016年15期)2016-03-01 02:46:28
山东医药(2015年32期)2016-01-20 13:53:50
现代制造技术与装备(2015年4期)2015-12-23 10:20:22
河北医科大学学报(2015年12期)2015-03-07 09:25:00
