
连续剪梢对金叶女贞不同叶位呈色和光合特性的影响
2013-08-02 00:52:10彭丽丽姜卫兵王明玉张斌斌
江苏农业学报 2013年2期
彭丽丽, 姜卫兵, 韩 健, 王明玉, 张斌斌
(1.南京农业大学园艺学院,江苏 南京 210095;2.江苏省农业科学院园艺研究所,江苏 南京 210014)
彩叶植物因其绚丽的叶色近年来在园林绿化中备受重视,应用越来越广泛[1]。金叶女贞(Ligustrum vicaryi L.)作为一种常见的彩叶植物,原分布于中国长江以南地区,现已引种到中国北方城市,在城市和园林绿化中发挥着重要作用[2]。金叶女贞单叶片的叶色是随着叶龄变化的,即幼叶呈金黄色(浅黄色),成熟叶呈黄绿色,老叶呈绿色。由于在同一枝条、同一植株上同时存在不同叶龄的叶片,所以同时呈现出金黄、黄绿、绿色等不同叶色的叶片,且自然生长情况下着生叶位具有规律性,即枝条上部(阳光直射处)叶片呈金黄色,枝条中部(树冠中部)叶片呈黄绿色,枝条下部(树冠内堂和基部)叶片呈绿色。
在城市和园林绿化管理中,整形修剪是一些彩叶树木栽培过程中重要养护措施之一,通过整形修剪可以满足观赏要求;整形修剪可促进衰老树、弱枝、弯曲枝更新复壮、加强树势,也可削弱过强枝长势以平衡枝条均衡分布;此外,整形修剪可改善树冠内通风透光条件[3-4]。目前关于修剪对彩叶树木光合生理的影响尚无报道,因此,本研究以2年生的盆栽金叶女贞为试验材料,研究剪梢条件下金叶女贞光合生理特性和色素含量的变化规律,以期为进一步完善金叶女贞的人工管理措施提供参考依据。
1 材料与方法
1.1 试验材料及处理
试验于2011年5月至9月在南京农业大学校园内进行。选用生长健壮,长势相近的2年生盆栽金叶女贞30株[种植于内径30 cm、深25 cm的花盆中,栽培基质为园土∶基质∶有机肥=1∶1∶1(体积比),每盆1株]。不剪梢(对照)和剪梢处理各15盆,采用常规管理,2011年5月摘除花序。
2011年5月中旬对剪梢处理组进行第1次剪梢处理(去除枝条上部金或浅黄叶),7月上旬和9月上旬分别对新枝条进行同样的处理。
1.2 方法
分别于2011年5月下旬、7月中下旬和9月中下旬的晴朗天气,选择不剪梢和剪梢金叶女贞南向的枝梢,测定中部功能叶、下部老叶光合色素含量、光合作用、叶绿素荧光参数,3次重复。
1.2.1 光合色素含量的测定 叶绿素(Chlorophyll,Chl.)和类胡萝卜素(Carotinoid,Car.)含量的测定采用张宪政[5]的方法。于8∶00,分别采摘植株中、下部叶位叶片,立即带回实验室,去掉主脉,剪碎混匀,称取0.2 g,用20 ml提取液(丙酮∶无水乙醇=1∶1,体积比)浸提24 h,并用保鲜膜封口防止提取液挥发,浸泡至组织变白后,用紫外/可见光分光光度计测定440 nm、645 nm、663 nm的OD值,叶绿素和类胡萝卜素含量的计算公式如下:叶绿素a(Ca,mg/L)=12.72A663- 2.59A645,叶绿素 b(Cb,mg/L)=22.88A645- 4.67A663,叶绿素(Ct,mg/L)=Ca+Cb,类胡萝卜素 (Cc,mg/L)=4.695A440-0.268(Ca+Cb),色素含量=(色素浓度×提取液体积×103)/叶片鲜重,单位为mg/g,重复3次,所有色素含量均以鲜重表示。
1.2.2 光合作用日变化的测定 选择晴朗无风天气,自7∶00~17∶00每隔2 h在完全模拟自然环境的条件下,选取完好的功能叶、老叶为测定对象,利用英国PP-System公司生产的CIRAS-1型光合测定系统测定叶片净光合速率(Pn)、蒸腾速率(Tr)等光合参数,每次重复3次,取平均值。水分利用效率WUE=Pn/Tr(取日平均值),利用AutoCAD软件计算日变化曲线围成的面积,得到Pn日积分值(Diurnal integral value of Pn,DIV of Pn)[6]。
1.2.3 叶绿素荧光参数的测定 采用FMS-2便携脉冲调制式荧光仪(英国Hansatech公司提供),与光合作用日变化同时进行测定叶片叶绿素荧光参数的日变化,测定的指标包括初始荧光(F0)、光系统Ⅱ(PSⅡ)最大光化学效率(Fv/Fm)、光系统Ⅱ(PSⅡ)实际光化学效率(Yield)。叶绿素荧光的光化学淬灭(qp)和非光化学淬灭(NPQ)方面的光能分配参照Quick等[7]提出的方法计算,电子传递速率ETR=0.50 ×0.85 × ΦPSⅡ × PAR[8]。各指标每次重复3次,均取日均值。
1.3 数据分析
利用Excel和Systatv7.0统计软件进行数据处理与分析。采用HSD法(P<0.05)进行差异显著性分析。
2 结果
2.1 连续剪梢对金叶女贞不同叶位叶片光合色素含量的影响
不剪梢(对照)与剪梢处理金叶女贞不同叶位光合色素含量(表1)显示:不同时期不剪梢处理(对照)金叶女贞中部叶位叶片叶绿素含量由大到小依次为:2011年9月、2011年7月、2011年5月,不同时期剪梢处理中部叶位叶片叶绿素含量由大到小依次为:2011年7月、2011年9月、2011年5月,不同时期不剪梢(对照)和剪梢处理下部叶位叶片叶绿素含量均由大到小依次为:2011年7月、2011年9月、2011年5月,差异均达到显著水平(P<0.05)。2011年7月剪梢处理中、下部叶位叶片叶绿素含量均显著大于不剪梢处理(对照)(P<0.05);2011年9月剪梢下部叶位叶片叶绿素含量显著大于不剪梢处理(对照)(P<0.05)。2011年5月不剪梢(对照)与剪梢处理金叶女贞中部叶位叶片类胡萝卜素含量均显著大于2011年7月、2011年9月(P<0.05);2011年7月不剪梢处理(对照)金叶女贞下部叶位叶片类胡萝卜素含量均显著大于2011年7月、2011年9月(P<0.05)。2011年5月剪梢处理下部叶位叶片类胡萝卜素含量显著大于不剪梢处理(对照)(P<0.05);2011年7月剪梢处理下部叶位叶片类胡萝卜素含量小于不剪梢处理(对照)(P>0.05),中部叶位叶片则相反。2011年5月不剪梢(对照)与剪梢处理中、下部叶位叶片叶绿素a/叶绿素b值均显著高于2011年7月和2011年9月(P<0.05);2011年5月和2011年7月不剪梢处理(对照)中、下部叶位叶片叶绿素a/叶绿素b值均高于2011年9月。2011年7月不剪梢(对照)与剪梢处理金叶女贞下部叶位叶片叶绿素/类胡萝卜素值均显著高于2011年5月和2011年9月;2011年7月剪梢处理下部叶位叶片叶绿素/类胡萝卜素值显著大于不剪梢处理(对照)(P<0.05)。
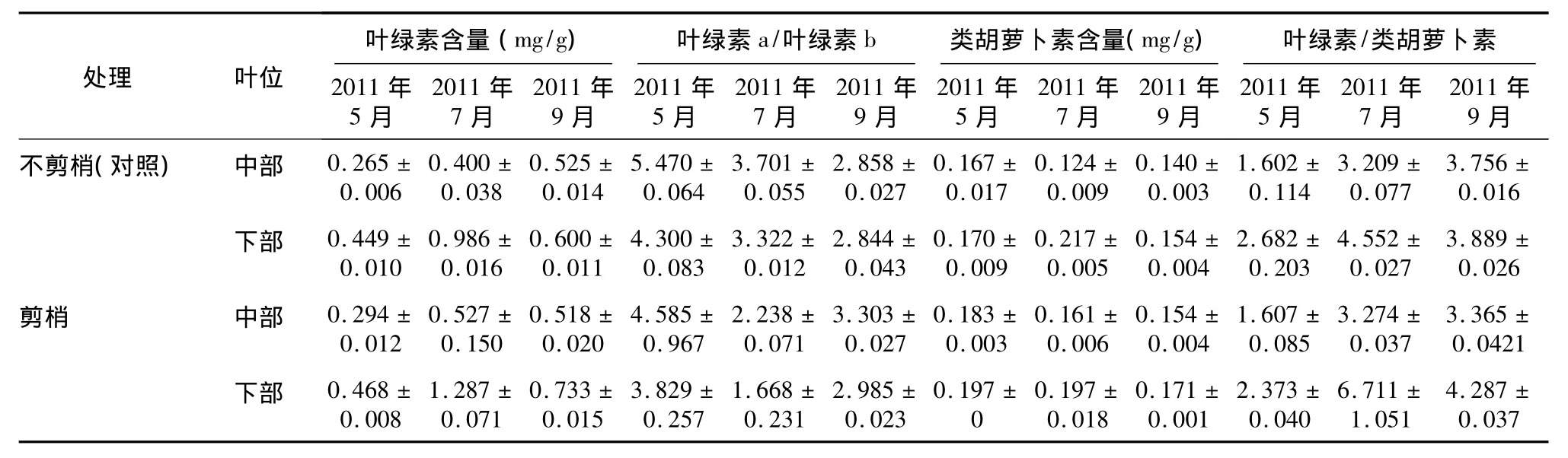
表1 剪梢对金叶女贞不同叶位叶片光合色素含量的影响Table 1 Effects of tip pruning on pigment content in middle and lower leaves of Ligustrum vicaryi
2.2 比较连续剪梢对不同叶位光合特性的影响
2.2.1 净光合速率(Pn)日变化与净光合速率日积分值(DIV of Pn) 图1表明,不同时期不剪梢(对照)与剪梢处理中部叶位叶片净光合速率日变化均呈双峰曲线,均在11∶00~13∶00间出现光合午休现象;不同时期不剪梢(对照)与剪梢处理下部叶位叶片净光合速率日变化均为单峰曲线。2011年7月剪梢处理金叶女贞中、下部叶片净光合速率均高于不剪梢处理(对照)。
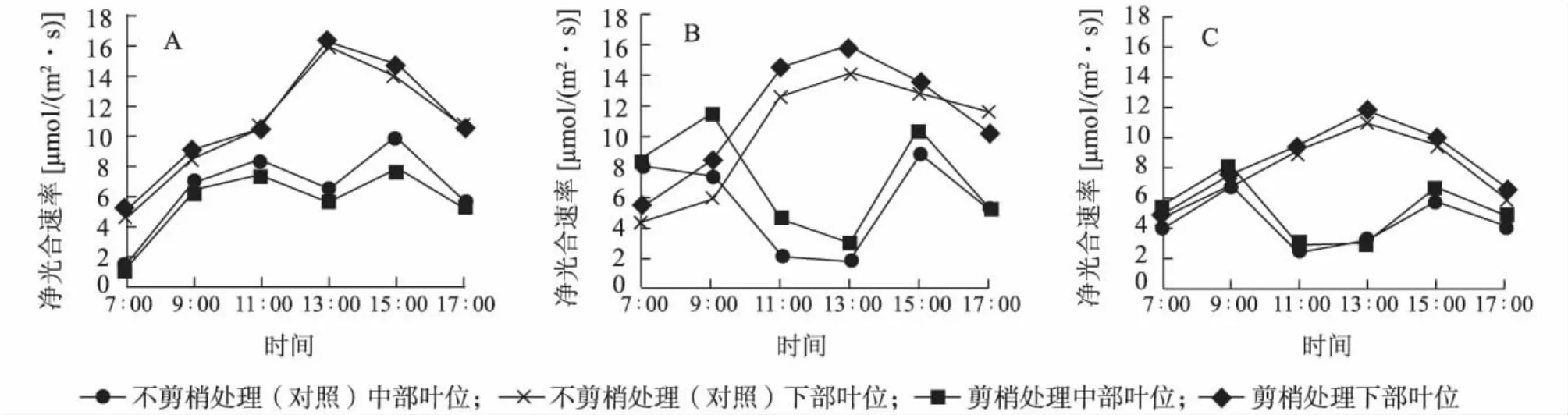
图1 净光合速率(Pn)日变化Fig.1 Diurnal variation of Pnin middle and lower leaves with or without pruning
净光合速率日积分值(DIV of Pn)可以反映一定时间内植物实际的光合净积累量。如图2所示,不剪梢处理(对照)金叶女贞中、下部叶位叶片净光合积累量随着时间的推进而下降。2011年7月剪梢处理金叶女贞中、下部叶位叶片净光合积累量大于2011年8月和2011年9月。
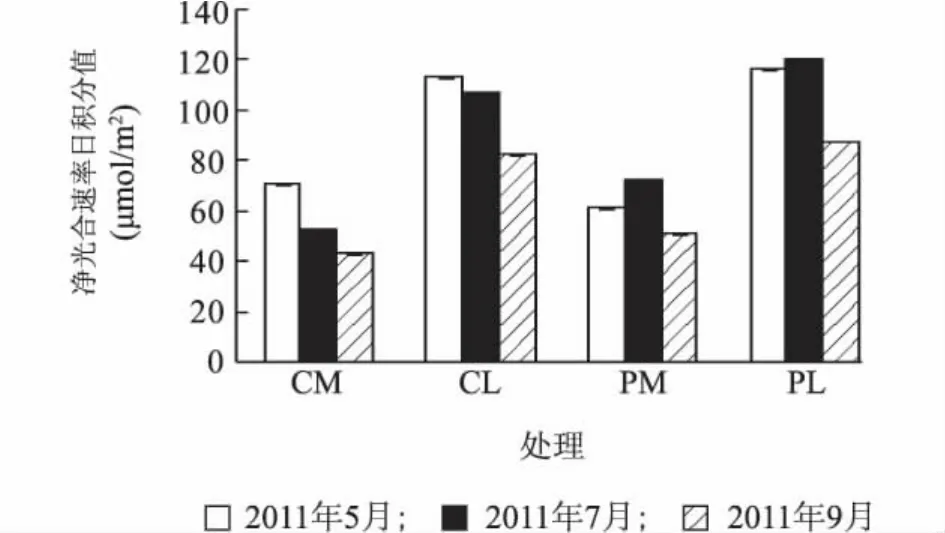
图2 不同叶位叶片净光合速率日积分值Fig.2 DIV of Pnand WUE of middle and lower leaves with or without pruning
2.2.2 水分利用效率(WUE)日变化 如图3所示,2011年7月、2011年9月白天上午随着温度光照强度的增加,不剪梢处理(对照)叶片水分利用效率降幅大于剪梢处理,自午间开始,二者变化趋势相近。

图3 水分利用效率(WUE)日变化Fig.3 Diurnal variation of WUE in middle and lower leaves with or without pruning
2.2.3 叶绿素荧光参数的比较 由图4可知,随着时间的推进,不剪梢处理(对照)中、下部叶位叶片的初始荧光(FO)随之增加。2011年5月、2011年7月剪梢处理中、下部叶位叶片F0均高于不剪梢处理(对照),2011年9月剪梢处理中、下部叶位叶片F0均低于不剪梢处理(对照)。不剪梢处理(对照)中、下部叶位叶片光系统Ⅱ(PSⅡ)最大光化学效率(Fv/Fm)变化趋势与F0一致。2011年5月、2011年7月剪梢处理中部叶位叶片Fv/Fm大于不剪梢处理(对照)(P<0.05)。2011年7月、2011年9月剪梢处理中部叶位叶片PSⅡ实际光化学效率(Yield)显著小于不剪梢处理(对照)(P<0.05),2011年9月剪梢处理下部叶位叶片Yield显著小于不剪梢处理(对照)(P<0.05)。不同时期剪梢处理下部叶位叶片qp均小于不剪梢处理(对照)。2011年7月、2011年9月剪梢处理中、下部叶位叶片非光化学猝灭(NPQ)均显著高于不剪梢处理(对照)(P<0.05)。不同时期剪梢处理中、下部叶位叶片电子传递速率(ETR)均低于不剪梢处理(对照)。

图4 不同叶位初始荧光(Fo)、PSⅡ最大光化学效率(Fv/Fm)、PSⅡ实际光化学效率(Yield)、电子传递速率(ETR)、叶绿素荧光的光化学淬灭(qp)和叶绿素荧光的非光化学淬灭(NPQ)Fig.4 Fo,Fv/Fm,Yield,ETR,qpand NPQ of middle and lower leaves with or without pruning
3 讨论
3.1 连续剪梢对金叶女贞光合特性的影响
前人研究认为,植物叶片在强光下难以全部消耗过剩光能,这些光能将产生光抑制,不利于植物叶绿素的生成,还会使其分解[9-10]。本研究结果显示,2011年9月剪梢处理中部叶片叶绿素含量小于不剪梢处理(对照),但2011年5月、2011年7月剪梢处理中、下部叶位叶片叶绿素含量均大于不剪梢处理(对照),这可能是因为剪梢改善中、下部叶位叶片的光照条件,同时也消除植株顶端优势,从而使物质分配发生转移,为叶绿素的合成提供了更多的底物。叶绿素含量的增加利于光合作用的进行[11-12]。本研究结果2011年7月剪梢处理金叶女贞中、下部叶片净光合速率均高于不剪梢处理(对照)。此外,2011年5月剪梢处理的中部叶位叶片光合速率降低,可能是中部叶位光合物质用于修复损伤[13]。
夏季植物接收太阳光能进行光合作用的同时也遭受强太阳辐射的胁迫,叶片光合机构吸收过剩光能后会引起开放PSII反应中心最大光化学效率(Fv/Fm)的降低,即产生光抑制现象,严重的光抑制可以导致反应中心的不可逆破坏[14]。2011年7月剪梢处理中部叶位叶片Fv/Fm和光化学猝灭系数(qp)均高于不剪梢处理(对照),这表明PSII反应中心的能量捕捉效率和转换效率增加,电子传递活性增大。2011年7月剪梢处理中部叶位叶片PSⅡ实际光化学效率(Yield)和电子传递速率(ETR)小于不剪梢处理(对照)。2011年7月剪梢和不剪梢(对照)处理净光合速率及叶绿素荧光参数(Fv/Fm、qp)结果表明,午间剪梢处理金叶女贞中部叶位叶片电子传递受抑制相对严重,加剧PSII反应中心的光抑制程度。
高等植物在系统发育过程中形成了对光合机构起重要保护作用的生理生化机制,这种机制依赖叶黄素循环和PSII反应中心可逆失活与热能耗散[15-16]。非光化学猝灭(NPQ)反映植物通过热耗散消耗过剩激能实现自身光保护的能力。本研究结果表明,连续剪梢处理在改善光合作用的同时激发了叶片光合机构的高速运转,更过多的过剩光能以热耗散形式散发,从而减轻伤害,这与前人的研究结果一致[17-19]。
3.2 连续剪梢对金叶女贞叶片光合色素含量的影响
金叶植物叶片在强光下叶色金黄,弱光下由于叶绿素含量增加,叶色转变为黄绿[20]。本研究结果表明,连续剪梢处理有利于金叶女贞中、下部叶位叶片类胡萝卜素的合成。2011年5月、2011年7月剪梢处理金叶女贞中、下部叶位叶片叶绿素/类胡萝卜素值高于不剪梢处理。表明完全剪除上部金黄叶或浅黄叶不利于金叶女贞叶片色彩的表达。
综上,2011年5月、2011年7月连续剪梢处理可促进金叶女贞叶片叶绿素的合成,有利于叶片光合作用,但2011年7月剪梢处理金叶女贞中部叶位叶片PSII反应中心午间的电子传递受阻更严重,光抑制程度加重。此外,完全去除上部金黄叶或浅黄叶不利于金叶女贞观赏价值的实现。因此,在园林绿化栽培修剪管理中,应该采取轻度修剪,保留部分金黄叶或浅黄叶以平衡其生长发育与观赏特性。
[1]姜卫兵,庄 猛,花国平,等.彩叶植物呈色机理及光合特性研究进展[J].园艺学报,2005,32(2):352-358.
[2]廖 容,崔 洁,卓春丽,等.成都市32种立体绿化植物降温增湿效应比较研究[J].江苏农业科学,2012,40(6):178-182.
[3]高文海,薛纯红.树木整形修剪的原因和季节[J].黑龙江科技信息,2009(7):269.
[4]黄冬贤,黎可华,黎明星,等.紫薇的整形修剪及园林应用[J].广东园林,2010(4):67-69.
[5]张宪政.作物生理研究法[M].北京:中国农业出版社,1986.
[6]庄 猛,姜卫兵,李志国.金边黄杨和大叶黄杨光合特性的比较[J].植物生理学通讯,2006,42(1):39-42.
[7]QUICK W P,STITT M.An examination of factors contributing to non-photochemical quenching of chlorophyll fluorescence in barley leaves[J].Biochimica and Biophysica Acta,1989,977:287-296.
[8]MAXWELLA K.JOHNSON G N.Chlorophyll fluorescence-a practical guide[J].Journal of Experimental Botany,2000,51(345):659-668.
[9]陆定志,傅家瑞,宋松泉.植物衰老及其调控[M].北京:中国农业出版社.1997.
[10]YANG X H,ZOU Q,WANG W.Photoinhibition in shaded cotton leaves after exposing to light and the time course of its restoration[J].Acta Botanica Sinica,2001,43:1255-1259.
[11]崔培强,姜卫兵,翁忙玲,等.遮荫对紫叶李幼苗叶片色素含量及光合速率的影响[J].西北植物学报,2010,30(11):2286-2292.
[12]姜 武,姜卫兵,李志国.园艺作物光合性状种质差异及遗传表现研究进展[J].经济林研究,2007,25(4):102-108.
[13]高桐梅,卫双玲,张海洋,等.打顶对芝麻不同叶位光合特性的影响[J].中国油料作物学报,2009,31(4):492-498.
[14]尤 鑫,龚吉蕊,段庆伟,等.两种杂交杨品系光合系统II叶绿素荧光特征[J].生态学报,2008,28(11):5641-5648.
[15]谢智华,姜卫兵,韩 键,等.叶片花色素苷对植物光合作用影响的研究进展[J].植物生理学报,2011,47(6):545-550.
[16]DEMMIG B,WINTER K,KROGER A,et al.Photoinhibit ion and zeaxanthin formation in int act leaves:A possible role of the xantho2 phyll cycle in the dissipat ion of excess light energy[J].Pl ant Physiol,1987,84:218-224.
[17]杨兴洪,邹 琦,赵世杰.遮荫和全光下生长的棉花光合作用和叶绿素荧光特征[J].植物生态学报,2005,29(1):8-15.
[18]YANG X,YI F,PRASAD P.Response of photosynthesis and chlorophyll fluorescence quenching to leaf dichotocarpism in Ligustrum vicaryi[J].Photosynthetica,2009,47(1):137-140.
[19]YANG X H,ZOU Q,WANG W.Photoinhibition in shaded cotton leaves after exposing to light and the time course of its restoration[J].Acta Botanica Sinica,2001(43):1255-1259.
[20]胡海姿,张 睿,尚爱芹,等.金叶植物色素含量对光强的响应[J].园艺学报,2007,34(3):717-722.
猜你喜欢
农业与技术(2023年11期)2023-06-19 02:28:18
读者(2023年2期)2023-02-20 08:10:40
干旱地区农业研究(2022年4期)2022-07-28 05:37:08
四川蚕业(2022年1期)2022-06-06 02:03:42
World Journal of Integrated Traditional and Western Medicine(2020年3期)2020-08-24 13:12:02
北京园林(2020年2期)2020-01-18 03:31:02
短篇小说(2018年4期)2018-05-21 08:27:15
基层中医药(2018年1期)2018-03-01 07:36:25
现代园艺(2017年11期)2017-06-28 11:32:18
河北林业科技(2016年5期)2016-11-08 03:12:56
