
砂砧薹草和假牛鞭草种群空间分布格局的分形特征
2013-07-24 01:49丁雪娇庞彩菊马玉心崔大练范彩彩
浙江农林大学学报 2013年2期
丁雪娇, 韩 红, 庞彩菊, 马玉心, 崔大练, 范彩彩
(1. 浙江海洋学院 海洋科学学院, 浙江 舟山316004; 2. 浙江农林大学 风景园林与建筑学院, 浙江 临安311300)
种群格局是种群生物学特性、 种内种间关系及环境综合作用的结果, 是种群基本数量特征之一[1]。种群格局的研究一直是生态学中研究的热点之一[2], 但目前仍有一些问题未能解决, 几乎共同存在的问题是种群格局类型依赖于尺度大小。 分形理论被认为是一个有用的解决这一问题的关键工具[3-4], 分形分析也强调了尺度的重要性, 尺度变化的内涵通过分形维数的变化反应出来[5]。 在生态学领域,应用分形维数能够客观地表达种群的分布式样, 同时也能反映种群占有空间、 利用资源的能力。 以分形几何分析种群格局多见于木本植物, 而草本植物研究的较少。 原因是草本群落分布较为集中, 尺度效应对其影响影响较小, 所以一般采用常规的样方分析方法。 对于海滨沙质海岸带, 受海水的侵蚀, 土壤异质性差异较大, 尺度的因素将直接影响种群结构的判别。 采用分形分析的方法将更准确地描述种群空间结构特征。 砂砧薹草Carex kobomugi 又名筛草, 莎草科Cyperaceae 多年生草本植物, 广泛分布黑龙江、 辽宁、吉林、 山东、 河北、 江苏、 浙江、 福建及台湾等沿海沙滩地区。 砂砧薹草是海滨重要的牧草植物, 其种子可以入药、 酿酒、 食用, 茎秆可以用于造纸。 假牛鞭草Parapholis incurva 为禾本科Poaceae 假牛鞭草属Parapholis 盐生杂草, 分布在热带、 亚热带和北半球的温带湿润地区, 也是重要的海滨牧草植物。 这2 种植物具有重要的生态价值, 均是海滨沙质沿岸的先锋植物, 能耐干旱瘠薄, 抗海风海雾, 还能适应海雾中夹杂的盐分胁迫和短期海潮造成的海浸。 它是改善沙岸滩涂生态环境, 防风固沙重要的植物资源[6]。笔者采用分形分析中的计盒维数和信息维数对分布于浙江舟山桃花岛海滨沙地的砂砧薹草和假牛鞭草种群个体空间关联的尺度变化特征进行研究, 以揭示出两者的种群空间分布格局的规律。
1 研究区概况
浙江舟山桃花岛位于舟山本岛南部, 29°46′~29°51′N, 122°13′~122°19′E, 岛长11.5 km, 宽约6.5 km, 面积为41.0 km2。 在桃花岛东侧的东海渔村具有很长的沙质海岸带, 旅游业称为“千步金沙”。 沙质海岸带长3 000 m 左右。 宽20~30 m。 沙质海岸平坦蜿蜓。 由于海陆热容量的显著差异, 形成了独特的海岛气候, 表现为四季分明、 冬暖夏凉、 雨水偏少、 台风频繁、 年日温差小、 以及最热月推迟至8 月、积雪稀少等。 由于该岛母岩大部分为凝灰岩, 也有少量砂页岩和花岗岩, 所以是海滨沙地形成的主要基质来源。 沙质盐土紧靠海边, 含盐量为10.0~15.0 g·kg-1, 是盐生植物主要生长区[7]。 植被分布主要有,肾叶打碗花Calystegia soldanella, 砂砧薹草, 假牛鞭草, 匍匐苦荬菜Ixeris repens, 珊瑚菜Glehnia littoralis, 单叶蔓荆Vitex trifolia var. simplicifolia, 绢毛飘拂草Fimbristylis sericea 等[8]。
2 研究方法
2.1 取样方法
在浙江舟山桃花岛海滨沙地选择具有代表性的地段作为调查样地。 由于海滨沙地呈带状分布, 样地的选择依据海滨地势, 选取长100 m 宽20 m 的带状样带。 在样带中随机取4 个砂砧薹草样方 (Q1~Q4),样方面积为1 m × 1 m; 随机取4 个假牛鞭草样方(Q5~Q8), 样方面积为1 m × 1 m。 在样方内采用相邻格子法取样, 对砂砧薹草和假牛鞭草进行定位观测记录, 并测量每个个体在样方平面坐标中的位置, 绘制点位图。
2.2 计算方法
2.2.1 计盒维数 计盒维数反映分形体对空间的占据程度, 取值一般是0~2。 计盒维数越大, 表明种群对空间的占据程度越大, 反之则越小[9]。 用格子边长为ε 的正方形网格对固定样地逐次进行栅格化处理,得到一系列不同网格边长为ε 时的非空格子数N(ε)。 计盒维数Db计算公式为:

根据野外绘制砂砧薹草和假牛鞭草点位图的分布情况, 用网格边长为0.5, 0.4, 0.3, 0.2, 0.1 m 的网格尺度, 对8 个样方分别进行网格栅格化处理, 计数非空格子数N(ε), 将每次划分所得的非空格子数N(ε)与对应的网格边长ε 在双对数直线坐标下进行直线拟合, 所得直线斜率的绝对值即为计盒维数估计值。
2.2.2 信息维数 种群格局的信息维数能反映格局强度的尺度变化。 信息维数高, 表明种群格局尺度变化强烈, 个体分布不均匀或聚集分布; 信息维数低, 表明种群格局强度尺度变化微弱, 个体均匀分布或随机性明显[10]。 信息维数Di的计算公式为:

根据野外绘制砂砧薹草和假牛鞭草点位图的分布情况, 用网格边长为0.5, 0.4, 0.3, 0.2, 0.1 m 的网格尺度, 对8 个样方分别进行网格栅格化处理。 在对种群分布格局的点位图进行网格覆盖的过程中, 进一步统计每个非空格中拥有的个体数量(Ni)。 若样地内的总个体数量为(N), 每个非空格中的个体分布概率为Pi=Ni/N, 信息量为Ii=-Piln(Pi), 则该网格边长时的总信息量为I(ε)=ΣIi。 将I(ε)与相应的网格边长ε 的对数值进行直线拟合, 得到的拟合直线斜率的绝对值即为信息维数估计值[11]。
3 结果与分析
3.1 计盒维数
计盒维数(box-counting dimension)能够揭示种群占据生态空间的程度和利用生态空间的能力。 在一定范围内存在的线性关系表明在该尺度范围内分维体对空间占据存在一致的规律[12]。
点位图是对种群分布的二维空间模拟, 其不仅能显示出种群个体在各样地的水平配置状况, 而且反映了砂砧薹草和不同样地的假牛鞭草种群在水平分布格局方面的差异[13]。 以此图为基础, 运用分形维数的计算方法, 可计算出砂砧薹草和假牛鞭草种群格局的计盒维数和信息维数, 进而揭示砂砧薹草和假牛鞭草个体空间关联、 系统结构复杂度和生态空间占据的多尺度变化规律[6]。 图1 为砂砧薹草和假牛鞭草种群分布格局位点图, 虽然从点位图中可以较直观地看出砂砧薹草和假牛鞭草种群在8 个样地内的分布格局, 但它只能粗略地估测出在样地中的分布特征。 从图2 可见: 在8 个样地中, lnε-lnN(ε)曲线图都存在2 个较明显的线性区域, 表明种群在2 个不同尺度范围内存在不同空间自相似性, 分别占据不同大小的生态空间, 说明计盒维数可反映砂砧薹草和假牛鞭草种群的分布格局。
通过直线拟合得到砂砧薹草和假牛鞭草种群的计盒维数(表1)。 由表1 可见: 8 个样地砂砧薹草和假牛鞭草种群的计盒维数分别为1.318 1, 1.596 5, 1.614 1, 1.511 1, 1.746 5, 1.639 6, 1.574 4 和1.454 2, 线性回归的相关系数都在0.950 0 以上, 相关极其显著(P<0.01), 表明回归直线的拟合程度很好, 砂砧薹草和假牛鞭草种群的空间格局具有明显的分形特征。 其中Q1~Q4为砂砧薹草种群, 其计盒维数为1.318 1~1.614 1; Q5~Q8为假牛鞭草种群, 其计盒维数为1.454 2~1.745 6。 从计盒维数的均值来看,假牛鞭草种群高于砂砧薹草, 说明其具有更强的占据空间的能力。 种群内的每个个体在其生长发育过程中都需要占据一定的空间资源, 由于每个个体的发育程度以及在群落空间中占据的位置不同, 其实际利用生态空间的能力也有所不同, 因此, 在不同群落中砂砧薹草和假牛鞭草种群的计盒维数存在差异。 其中: 样地3, 样地5 和样地6 的分株种群个体数量较多且在样地中分布面积较广, 种群表现出较强的空间占据能力, 其格局的计盒维数较大, 而样地1 和样地8 的分株种群数量相对较少且呈数个斑块状分布, 占据空间的能力较弱, 种群格局的计盒维数较小。 Q1~Q4砂砧薹草种群的计盒维数大小依次为Q3>Q2>Q4>Q1, Q5~Q8假牛鞭草种群的计盒维数大小依次为Q5>Q6>Q7>Q8, 8 个样方中砂砧薹草和假牛鞭草种群的计盒维数大小依次为Q5>Q6>Q3>Q2>Q7>Q4>Q8>Q1。
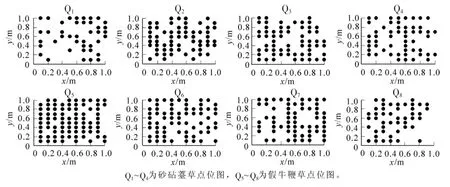
图1 砂砧薹草和假牛鞭草种群分布格局位点图Figure 1 Spatial distribution patterns of eight Carex kobomugi and Parapholis incurva populations
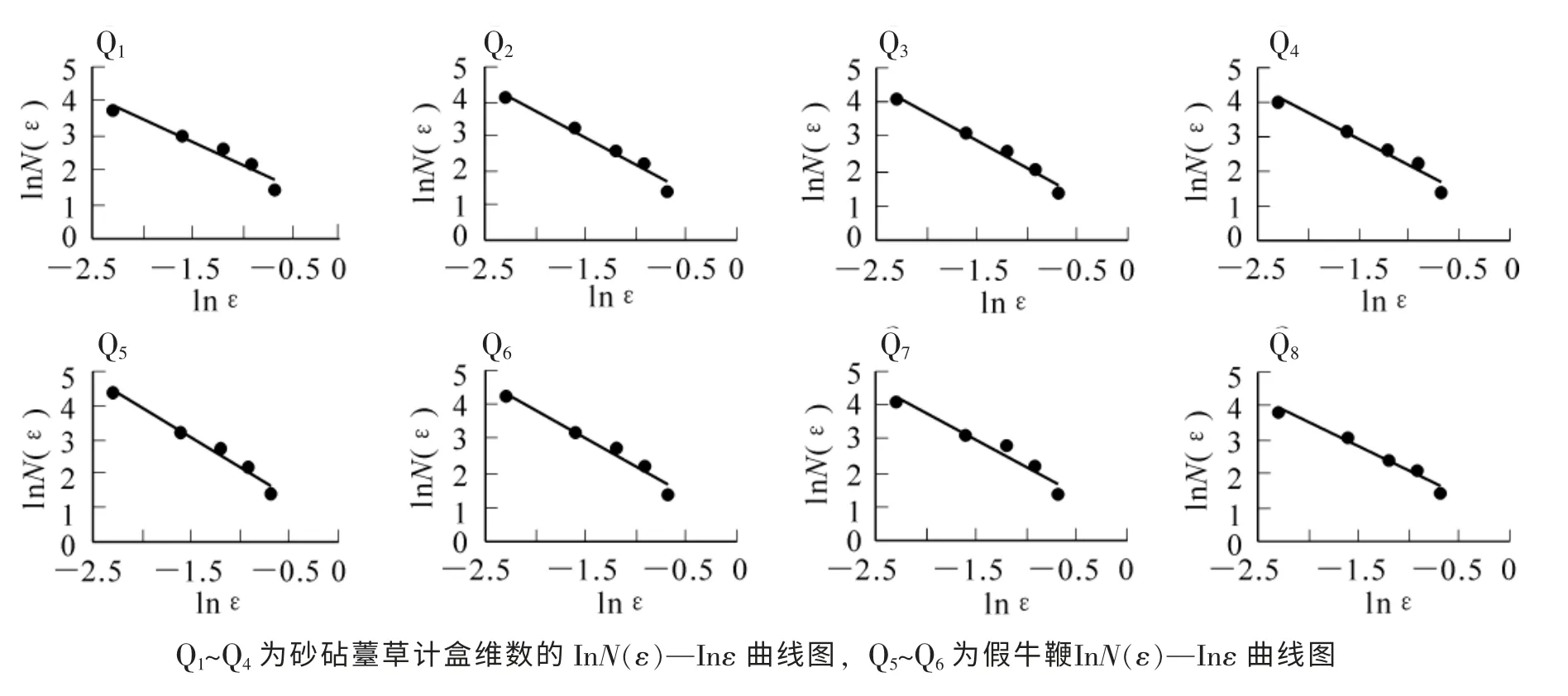
图2 砂砧薹草和假牛鞭草种群格局计盒维数的lnN(ε)—lnε 曲线图Figure 2 The lnN(ε)—lnε curves of the box-counting dimension of Carex kobomugi and Parapholis incurva population
3.2 信息维数
信息维数(information dimension)表征了不同尺度上系统结构复杂性之间的联系和规律性, 揭示了系统结构复杂性的尺度变化程度。 它是计盒维数的推广, 其不但考虑所取格子是“空”还是“非空”, 而且考虑了不同的非空格子所提供的信息量大小, 从而反映种群空间分布的非均匀性程度。 高的信息维数(接近2)表明种群格局强度尺度变化强烈, 个体分布不均匀, 随着观测尺度的改变, 种群格局分布类型会出现明显的变化[7]。
根据表2, 格子边长对数值lnε 与总信息量I(ε)之间的直线拟合的相关系数均在0.950 0 以上, 线性关系明显, 达到极显著的相关水平(P<0.01), 表明直线回归的斜率能很好地表征克隆种群空间结构的信息维数。 本研究中计算的8 块样地中信息维数分别为1.027 0, 1.257 3, 1.263 4, 1.209 4, 1.284 5,1.381 2, 1.246 4, 1.022 7(图3)。 其中Q1~Q4为砂砧薹草种群, 其信息维数为1.027 0~1.263 4; Q5~Q8为假牛鞭草种群, 其信息维数为1.022 7~1.381 2。 从信息维数的均值来看, 假牛鞭草种群高于砂砧薹草, 说明假牛鞭草种群比砂砧薹草种群更复杂。 通常, 如果种群聚集成块, 个体分布不均匀, 会导致信息维数较高, 反之, 个体星散分布, 随机性明显, 或个体均匀分布, 格局强度较低, 则信息维数较低。因此, 信息维数的差异亦揭示了种群更新状况的差异。 由此从数据中可以看出: 假牛鞭草种群的格局强度及聚集强度明显高于砂砧薹草。
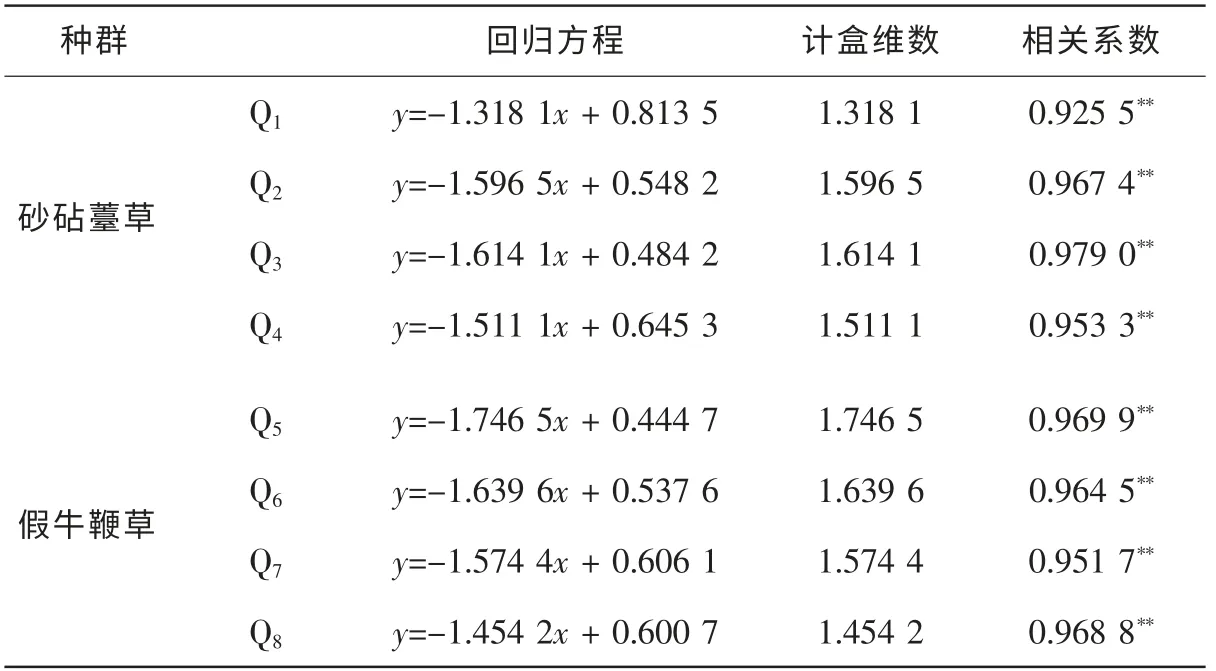
表1 砂砧薹草和假牛鞭草种群格局的计盒维数Table 1 Box-counting dimensions of the distribution patterns of eight Carex kobomugi and Parapholis incurva populations
4 结论与讨论
分形理论主要研究某一测度指标随尺度变化的自相似规律。 分形维数是描述这种自相似规律的指标,常用的是计盒维数、 信息维数和关联维数。 分形维数高, 则表明物种占据空间的能力强, 物种能更好地利用资源, 在与其他物种的竞争中处于优势地位。 假牛鞭草的计盒维数及信息维数的均值高于砂砧薹草, 说明假牛鞭草种群占据空间的能力比砂砧薹草强, 也说明假牛鞭草种群的格局强度高于砂砧薹草。
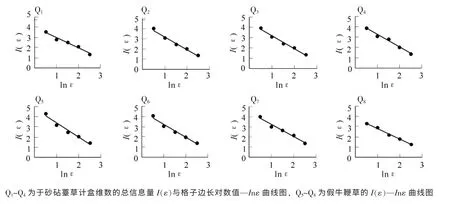
图3 砂砧薹草和假牛鞭草种群格局信息维数的I(ε)—ln(ε)曲线图Figure 3 I(ε)—ln(ε) curves of the information dimension of Carex kobomugi and Parapholis incurva population
植物种群分布格局分形特征方面的研究多集中在对木本植物的研究上, 而关于草本植物的分形特征研究较少。 倪红伟等[14]应用分形理论对草本植物小叶章Deyeuxia angustifolia 种群的分形特征进行了研究, 其结果说明克隆植物小叶章种群的分布格局是具有分形特征的。从本研究可知: 克隆植物砂砧薹草及假牛鞭草种群空间结构也具有分形特征。 这就更加证明, 分形理论在研究草本植物种群空间结构中具有广阔的前景。
计盒维数与信息维数与植物的克隆指数密切相关。 基株克隆构型变化与种群生态空间占据能力密切相关, 基株克隆构型趋于密集型的种群具有较强的空间占据能力, 而基株构型趋于游击型的种群, 其空间占据能力较弱[12]。 砂砧薹草与假牛便草克隆构型不同, 砂砧薹草其根状茎垂直向下为40~50 cm, 其克隆主要依靠以根状茎分枝形成。 而假牛鞭草自基部第2 节即具有分枝, 而且其分枝沿地面匍匐, 这2种植物均是克隆植物, 可能是假牛鞭草具有更强的克隆指数, 假牛鞭草具有地上的匍匐茎而砂砧薹草没有, 其地上匍匐茎在地面蔓延很长, 其对空间的占据能力可能更强。 砂砧薹草种群, 其计盒维数为1.318 1~1.614 1, 假牛鞭草种群, 其计盒维数为1.454 2~1.745 6, 其均值大于1.500 0, 说明对空间具有较强的占据能力, 而且假牛鞭草比砂砧薹草具有更强的占据空间的能力。
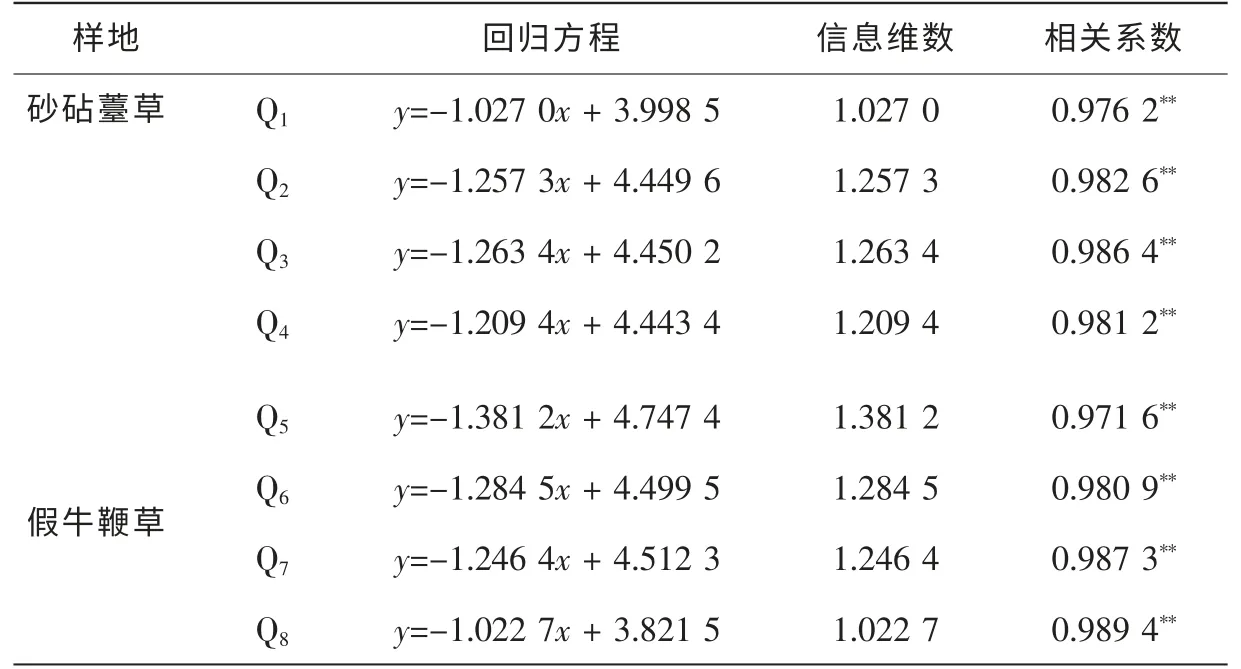
表2 砂砧薹草和假牛鞭草种群格局的信息维数Table 2 Information dimensions of the distribution patterns of eightCarex kobomugi and Parapholis incurva populations
总之, 砂砧薹草种群与假牛鞭草种群的计盒维数均值大于1.500 0, 具有较强的占据空间的能力,而且假牛鞭草种群由于具有更强的克隆构型, 其计盒维数大于砂砧薹草, 具有更强的占据空间的能力。假牛鞭草的信息维数均值大于砂砧薹草, 所以其种群的格局强度及群聚度更大, 种群结构更加复杂。
[1] 曲仲湘, 吴玉树, 王焕校, 等. 植物生态学[M]. 2 版. 北京: 高等教育出版社, 1983.
[2] 王本洋, 余世孝. 种群分布格局的多尺度分析[J]. 植物生态学报, 2005, 29 (2): 235-241.WANG Benyang, YU Shixiao. Multi-scale analyses of population distribution [J]. Acta Phytoecol Sin, 2005, 29 (2):235-241.
[3] 马克明, 祖元刚. 植被格局的分形特征[J]. 植物生态学报, 2000, 24 (1): 111-117.MA Keming, ZU Yuangang.Fractal properties of vegetation pattern [J].Acta Phytoecol Sin, 2000, 24 (1): 111-117.
[4] 宋萍, 洪伟, 吴承祯, 等. 天然黄山松种群空间格局的分形特征——关联维数[J]. 武汉植物学研究, 2005, 3(1): 32-36.SONG Ping, HONG Wei, WU Chengzhen, et al. Fractal characteristics of spatial pattern of natural Pinus taiwanens is population: correlation dimension [J]. J Wuhan Bot Res, 2005, 3 (1): 32-36.
[5] 戴小华, 余世孝. GIS 支持下的种群分布格局分析[J]. 中山大学学报: 自然科学版, 2003, 42 (1): 75-78.DAI Xiaohua, YU Shixiao. Analysis of population distribution pattern based on GIS technique [J]. Acta Sci Nat Univ Sunyatseni, 2003, 42 (1): 75-78.
[6] 乔勇进, 谢韶颖. 沙质海岸的先锋植物: 砂钻薹草[J]. 植物杂志, 2003 (4): 12.QIAO Yongjin, XIE Shaoying. The pioneer plant of the sandy coast: Carex kobomugi [J]. Plant J, 2003 (4): 12.
[7] 郭华, 王孝安, 肖娅萍. 秦岭太白红杉种群空间分布格局动态及分形特征研究[J]. 应用生态学报, 2005, 16(2): 227-232.GUO Hua, WANG Xiao’an, XIAO Yaping. Spatial distribution pattern and fractal analysis of Larix chinensis populations in Qinling Mountain [J]. Chin J Appl Ecol, 2005, 16 (2): 227-232.
[8] 李根有, 周世良, 张若蕙, 等. 浙江舟山桃花岛的天然植被类型[J]. 浙江林学院学报, 1989, 6 (3): 243-254.LI Genyou, ZHOU Shiliang, ZHANG Ruohui, et al. Zhejiang Zhoushan Taohua Island natural vegetation types [J]. J Zhejiang For Coll, 1989, 6 (3): 243-254.
[9] 田玉强, 李新, 江明喜. 后河自然保护区珍稀濒危植物种群分布格局的分形特征: 计盒维数[J]. 应用生态学报,2003, 14 (5): 681-684.TIAN Yuqiang, LI Xin, JIANG Mingxi. Fractal properties of the spatial pattern of rare and endangered plant populations in Houhe Nature Reserve in Hubei: box-counting dimension [J]. J Appl Ecol, 2003, 14 (5): 681-684.
[10] 石胜友, 成明昊, 郭启高. 涪陵磨盘沟桫椤种群格局的分形特征-信息维数[J]. 西北植物学报, 2004, 24 (7):1179-1183.SHI Shengyou, CHENG Minghao, GUO Qigao. Information dimension fractal properties of the spatial pattern of Alsophila spinulosa population in Mopangou, Fuling [J].Acta Bot Boreali-Occident Sin, 2004, 24 (7): 1179-1183.
[11] 马克明, 祖元刚. 兴安落叶松种群格局的分形特征: 信息维数[J]. 生态学报, 2000, 14 (5): 681-684.MA Keming, ZU Yuangang. Fractal properties of the spatial pattern of Larix gmelini population: information dimension [J]. Acta Ecol Sin, 2000, 14 (5): 681-684.
[12] 向悟生, 李先棍, 苏宗明, 等. 元宝山南方红豆杉克隆种群分布格局的分形特征[J]. 植物生态学报, 2007, 31(4): 568-575.XIANG Wusheng, LI Xiangun, SU Zongming, et al. Fractal properties of distribution patterns of Taxus chinensis var. mairei clonal populations in the Yuanbaoshan Mountain, China [J]. J Plant Ecol, 2007, 31 (4): 568-575.
[13] 林勇明, 吴承祯, 洪伟, 等. 香料植物桂花野生种群分布格局的分形分析[J]. 中国农学通报, 2009, 25 (20):121-124.LIN Yongming, WU Chengzhen, HONG Wei, et al. Fractal properties of distribution pattern of natural Osmanthus fragrans populations [J]. Chin Agric Sci Bull, 2009, 25 (20): 121-124.
[14] 倪红伟, 郭亚胜, 吴海一. 小叶章种群分布格局的分形特征Ⅱ信息维数与关联维数[J]. 木本植物研究, 2000, 20(3): 351-354.NI Hongwei, GUO Yasheng, WU Haiyi. The fractal character of the distribution pattern of Deyeuxia angustifolia population (Ⅱ) information dimension and correlation dimension [J]. Bull Bot Res, 2000, 20 (3): 351-354.
猜你喜欢
闽南师范大学学报(自然科学版)(2022年3期)2022-12-06
湖北大学学报(自然科学版)(2022年3期)2022-12-01
东坡赤壁诗词(2021年1期)2021-03-24
延安大学学报(自然科学版)(2020年4期)2021-01-15
动漫星空(兴趣百科)(2020年11期)2020-11-09
东坡赤壁诗词(2020年4期)2020-09-02
中国药房(2019年24期)2019-09-10
趣味(数学)(2019年12期)2019-04-13
现代装饰(2018年11期)2018-11-22
