
毛竹林下多花黄精种群生长和生物量分配的立竹密度效应
2013-07-24 01:30樊艳荣陈双林杨清平李迎春郭子武
浙江农林大学学报 2013年2期
樊艳荣, 陈双林, 杨清平, 李迎春, 郭子武, 陈 珊
(中国林业科学研究院 亚热带林业研究所, 浙江 富阳311400)
毛竹Phyllostachys edulis 是中国最为著名的经济竹种, 分布面广, 利用价值高, 经济效益好, 对区域农村社会经济贡献率大。 然而, 散生竹开花结实困难, 遗传育种难度大, 提高单产主要依赖于林分结构调控和土壤管理, 而集约经营毛竹林进一步提高竹材、 竹笋产量的潜力空间已越来越小, 还面临着立地生产力衰退, 及农村劳动力、 农资成本不断上升, 经济效益明显下滑等问题。 鉴于毛竹林土地空间资源利用率低, 植物种间生态位互补特征等, 可以实行复合经营来解决上述问题。 目前, 已开展了一些竹林复合经营的研究与实践, 主要集中在毛竹与杉木Cunninghamia lanceolata[1-5], 马尾松Pinus massoniana[6-7],阔叶树[8-9]等的混交经营, 表明实行竹木混交经营, 可以充分利用营养空间, 改善土壤养分状况, 提高林地生产力和竹林产品质量, 有利于毛竹林可持续经营。 而就毛竹与药用植物的复合经营研究很少涉及。 多花黄精Polygonatum cyrtonema 隶属百合科Liliaceae 黄精属Polygonatum, 分布区域广, 是药食同源的多年生草本植物, 中国传统大宗药材, 利用部分为横生、 肥厚的地下块茎, 具有补气养阴、 健脾、润肺、 益肾等功效, 是工厂化生产药品、 保健品等的原料。 近年来陆续开展了较多对多花黄精的相关研究, 主要集中于地下块茎化学成分的提取和活性鉴定[10-11]及形态指标观测[12-14]等, 而对于多花黄精在林下的复合栽培研究很少涉及。 根据多花黄精是人工粗放经营毛竹林中的常见种, 从理论上分析可以实行毛竹-多花黄精复合经营。 生物量及其分配格局受气候、 地形、 光照和土壤水分、 养分等生态因子的深刻影响, 林分密度通过影响林内的光照、 温度、 湿度等环境条件, 对植物群落的生物量在层片、 器官水平上的分配起到重要的影响作用[15]。 立竹密度是否会明显影响到毛竹林下多花黄精的生长和生物量分配格局, 尤其是药食同源利用部分的地下块茎生长和生物量产出呢? 为此, 本研究以人工粗放经营的毛竹林为对象, 通过立地条件基本一致的不同立竹密度毛竹林下多花黄精种群生长和生物量积累与分配格局的研究, 试图明确毛竹林下多花黄精能够良好生长的适宜立竹密度, 这对于毛竹-多花黄精复合经营理论的建立和技术的形成非常重要。
1 研究地概况与方法
1.1 研究区自然概况
试验地位于浙江省江山市(28°15′26″~28°53′27″N, 118°22′37″~118°48′48″E )保安乡, 处于浙、 闽、赣三省交界处。 气候温暖湿润, 四季分明, 雨水充沛, 年降水量为1 650~2 200 mm, 相对湿度85%~95%, 年平均气温17.1 ℃, 年均日照时数2 063.3 h, 年均无霜期253 d。 土壤为红壤, 厚度1 m 以上。试验地毛竹林资源丰富, 面积800 hm2, 是江山市毛竹主要产出乡镇, 但竹林经营粗放, 毛竹林多为纯林, 以材用林经营为主, 实行季节性伐竹和留笋养竹, 没有化学除草剂使用历史, 未采取过林地垦复和施肥等措施。
1.2 试验毛竹林设置
由于试验区不同经营户的毛竹林伐竹、 留笋养竹强度不同, 使林分立竹密度存在差异。 根据试验区材用毛竹林立竹密度状况, 试验选择了3 种立竹密度的毛竹林, 立竹密度分别为D1(1 500~2 500 株·hm-2),D2(2 500~3 500 株·hm-2), D3(3 500~4 500 株·hm-2), 其中, D2密度是毛竹材用纯林丰产经营适宜的立竹密度。 D1, D2和D3试验林所在海拔、 坡度、 坡长分别为502 m, 30~34 度, 110 m; 505 m, 31~35 度, 80 m; 511 m, 29~34 度, 50 m。 均为下坡位, 坡向均为半阴坡。 由于试验毛竹林海拔相近, 坡位、 坡向相同, 可以认为立地条件基本一致。 根据3 种立竹密度试验毛竹林20 m × 20 m 样地的林分结构调查数据统计, 试验毛竹林林分结构见表1。

表1 试验毛竹林林分结构Table 1 Stand structures of Phyllostachys edulis forest
1.3 试验毛竹林地土壤化学性质
分别在3 种立竹密度试验毛竹林中随机布点(3 点) 挖取土壤剖面(0~30 cm)取土样, 将3 个点的土样混合均匀, 取土样1.0 kg 左右, 重复3 次。 全氮用凯氏法测定, 全磷用酸溶-钼锑抗比色法测定, 全钾用酸溶火焰光度法测定, 有机质用重铬酸钾容量法测定, 速效氮用碱解法测定, 速效磷用盐酸氟化铵法提取钼锑抗比色法测定, 速效钾用乙酸铵浸提火焰光度法测定, pH 值用酸度计测定[16]。 试验毛竹林土壤化学性质分析结果见表2。
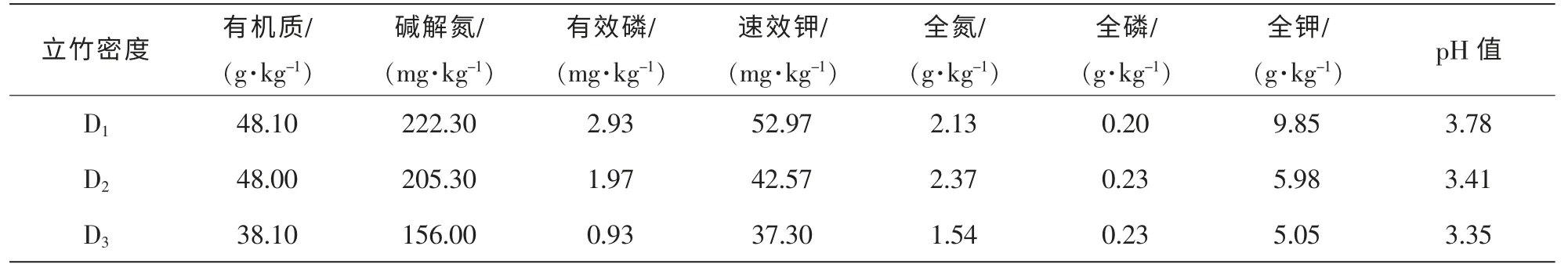
表2 试验毛竹林土壤化学性质Table 2 Soil chemical properties of Phyllostachys edulis forest
1.4 试验调查方法
2011 年8 月在不同立竹密度的试验毛竹林中各设置3 个20 m × 20 m 样地(样地间相隔20 m 以上,土壤厚度基本一致), 沿等高线在每个样地的上部、 中部、 下部各设置3 个2 m × 2 m 的样方, 设样方9个·样地-1。 对每个样方测量每株多花黄精的地径和株高, 对每个样地的立竹进行胸径测量, 统计立竹密度。 另在3 种立竹密度的试验毛竹林中各随机选取20 株多花黄精样株, 整株挖起, 分离叶、 地上茎、根、 地下块茎, 分别称鲜质量, 然后置于80 ℃烘箱中烘至恒量, 称干质量, 分析各构件生物量分配比例。
叶片SPAD 值(specialty products agricultural division)为叶绿素相对值(chlorophyll relative value), 是植物叶绿素相对含量的读数, 也称绿色度[17], 测定方便, 已在农林业科学研究中得到了较为广泛的应用[18-19],所以本研究也用叶片SPAD 值指标来分析多花黄精的生长状况。 具体测定方法为随机选取多花黄精样株3 株·样方-1, 分别在冠层的上部、 中部、 下部各选取1 片生长正常的叶片用德国产的SPAD-502 叶绿素测定仪(Minolta SPAD-502 chlorophyll meter)测定叶片SPAD 值, 3 次重复。
1.5 数据处理与分析
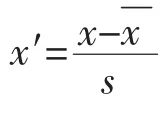
2 结果与分析
2.1 毛竹林立竹密度对多花黄精生长的影响
毛竹林立竹密度对多花黄精的地径、 叶片叶绿素值无显著影响, 而对多花黄精的种群密度、 株高有一定的影响, 均随着立竹密度的增大而减小(表3), 且D1立竹密度毛竹林的多花黄精种群密度、 株高显著地高于D2和D3立竹密度毛竹林(P<0.05), 其中, 种群密度分别增加了79.17%和95.45%, 株高分别提高了34.63%和72.50%, 而D2和D3立竹密度毛竹林的多花黄精种群密度、 株高无显著差异(P>0.05)。
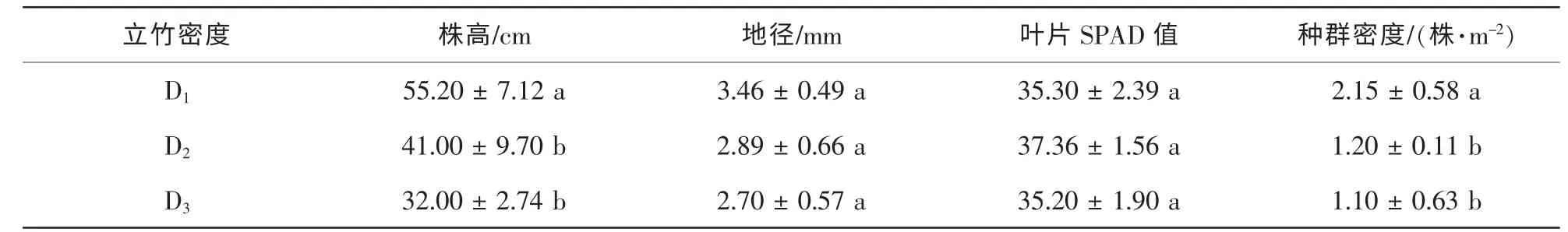
表3 不同立竹密度毛竹林多花黄精生长状况Table 3 Growth conditions of Polygonatum cyrtonema in Phyllostachys edulis forest at different bamboo densities
2.2 毛竹林立竹密度对多花黄精种群生物量积累和分配的影响
毛竹林立竹密度对单位面积的多花黄精各构件生物量和总生物量积累均有一定的影响, 且多花黄精各构件生物量和总生物量均随着立竹密度的增大而减小(图1)。 多花黄精叶生物量D1立竹密度毛竹林显著高于D3立竹密度毛竹林(P<0.05), 但与D2立竹密度毛竹林差异不显著(P>0.05); 地上茎、 根生物量均是D1立竹密度毛竹林显著地高于D2和D3立竹密度毛竹林(P<0.05), 而后两者无显著差异(P>0.05);地下块茎和总生物量D1, D2和D3立竹密度毛竹林间均有显著差异(P<0.05)。
从表4 分析可知: 不同立竹密度毛竹林下多花黄精相同构件的生物量分配比例均无显著差异(P>0.05), 多花黄精构件生物量分配格局均为地下块茎>根、 地上茎、 叶, 地下块茎生物量所占比例显著大于其他器官(P<0.05), 达70%~79%,而D1, D2和D3立竹密度毛竹林下多花黄精的叶、 地上茎、 根生物量分配比例间均无显著差异(P>0.05), 说明多花黄精除地下块茎外的其他器官生物量分配较为均匀。
2.3 毛竹林多花黄精种群生长及生物量综合指标的因子分析
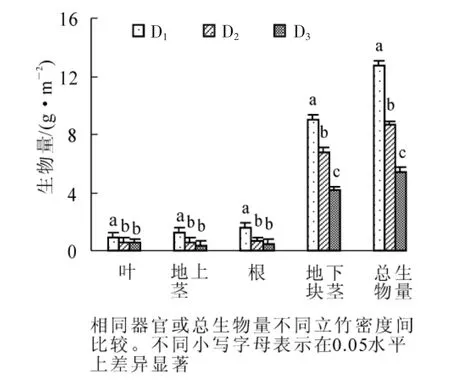
图1 不同立竹密度毛竹林多花黄精生物量积累Figure 1 Biomass accumulation of Polygonatum cyrtonema in Phyllostachys edulis forest at different bamboo density
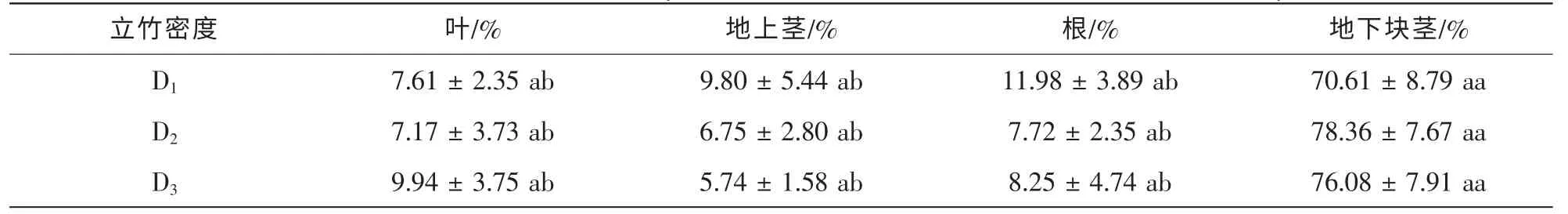
表4 不同立竹密度毛竹林多花黄精生物量分配Table 4 Biomass allocation of P. cyrtonema in Ph. edulis forest at different bamboo density
对多花黄精生长和构件生物量指标进行因子分析, 其中对于公因子的提取运用主成分分析,得到主成分的特征值和贡献率(表5), 其中: 前2 个主成分包含了多花黄精生长和生物量的全部信息,因此提取前2 个主成分作为描述多花黄精生长和生物量的综合指标。
根据特征值可求出各主成分的特征向量, 进而求出主成分方程, 前2 个公因子方程为: y1=0.991x1+1.000x2-0.218x3+0.984x4+0.994x5+1.000x6+0.995x7+0.950x8+0.983x9; y2=0.131x1-0.021x2+0.976x3-0.176x4-0.109x5-0.096x7+0.313x8+0.186x9。 其中: y 代表公因子, x1~x9依次代表多花黄精株高、 地径、 叶片叶绿素值、 种群密度、 叶生物量、 地上茎生物量、 根生物量、 地下块茎生物量、 总生物量。
第1 公因子代表了87%以上的信息, 在全部变量中处于最重要的地位。 其中: 株高、 地径、 种群密度、 叶生物量、 地上茎生物量、 根生物量、 地下块茎生物量、 总生物量等指标的特征向量系数较大, 因此可以将第1 公因子看作是表征多花黄精生长及生物量的综合指标。
第2 公因子代表了近13%的信息。 其中: 叶片叶绿素值的特征向量系数较大, 因此, 可以将第2 公因子看作是表征多花黄精叶片叶绿素值的指标。
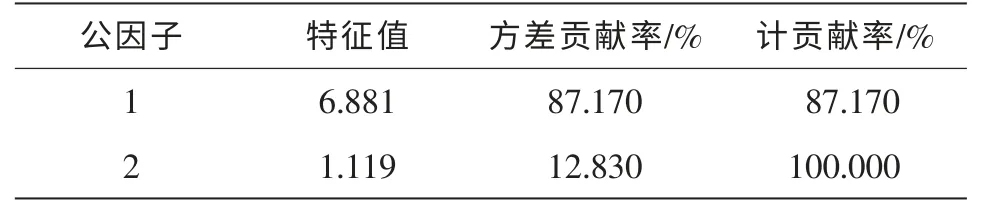
表5 各公因子特征值和方差贡献率Table 5 Principal components’ eigenvalue and variance contribution
2.4 毛竹林多花黄精种群生长及生物量指标的综合比较
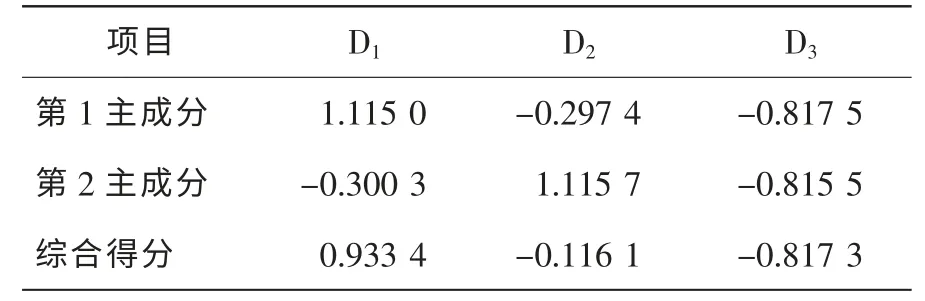
表6 毛竹林下多花黄精生长和生物量指标主成分综合得分Table 6 Principal component comprehensive scores of P.cyrtonema’s growth and biomass in Ph.edulis forest
3 结论与讨论
林分密度对林下植被生长和生物量分配等的影响机制是多方面的, 它可以通过改变林分内的温度、湿度等坏境条件来限制林下植被的生长, 但主要通过改变林分中的光资源来影响, 获光率是影响林下植被生存和生长的潜在因素, 林内光照的改变将会影响到林下植被的种类、 数量和生物量分布等[20]。 本研究表明: 毛竹林立竹密度是影响多花黄精种群生长的重要因素, 以D1立竹密度最好, D3立竹密度最差,这与随着毛竹林立竹密度的增大, 林内光照变弱, 多花黄精植株得不到充分的光照从而影响其生长乃至生存有着密切的关系。 试验毛竹林林分结构的差异是由人为留笋养竹和伐竹引起的, 立地条件基本一致。 D3试验林不仅立竹密度较D1和D2试验林大, 而且立竹胸径也较D1和D2试验林明显增大, 意味着立竹冠幅明显增大, 使林分郁闭程度显著提高, 林内光照强度明显减弱, 说明林内光照对多花黄精的生长有着重要的影响。 这也可从多花黄精裸地(不遮光)和透光率分别为25%, 40%, 55%的遮阳栽培处理结果为40%透光率遮阳处理时多花黄精生长势最强[21]中得到体现, 光照强度过大和过小均不利于多花黄精生长。 试验表明D2试验林立竹密度是多花黄精种群生长显著变化的“节点”, 是毛竹-多花黄精复合经营的立竹密度上限。
植物生物量的积累和分配状况在很大程度上反映了植物对光能、 水分和养分的利用效率。 地上部分生物量的合理配置是植物对生长环境长期适应的结果, 能增加植物对光能的利用效率, 促进有机物的积累, 而地下部分生物量的分配状况则体现了植物对土壤水分和无机养分的吸收能力[22]。 本研究表明: 毛竹林立竹密度也是影响多花黄精种群生物量积累的重要因素, 以D1立竹密度最好, D3立竹密度最差,D3立竹密度毛竹林下多花黄精地下块茎生物量和总生物量积累均显著低于D2立竹密度毛竹林(P<0.05),而种群密度、 地径、 株高和叶片SPAD 值所反映的种群生长状况差异不明显, 说明林内光照对多花黄精目标栽培的地下块茎生长和生物量积累有着重要的影响, 这也可从多花黄精露地栽培、 苹果林套种为后者多花黄精地下块茎增产效果好[21]中得到反映。
不同立竹密度毛竹林下多花黄精相同构件的生物量分配比例均无显著差异(P>0.05), 多花黄精构件生物量分配格局均为地下块茎>根、 地上茎、 叶, 地下块茎生物量所占比例显著大于生物量分配较为均匀的其他器官(P<0.05), 达70%~79%。 多花黄精把大量的生物量分配给地下系统(块茎和根) , 有利于充分利用土壤养分空间, 增加土壤养分、 水分的吸收效率, 并能够有效地保存, 有利于种群的维持和扩大, 这对于多花黄精种群生长和生存有着积极的生态学意义。 根据主成分R 型因子综合评定法, 不同立竹密度毛竹林下多花黄精生长和生物量特征综合得分大小顺序为D1>D2>D3, 结合药食同源利用的器官(地下块茎)生物量积累也是D1立竹密度毛竹林显著大于D2和D3立竹密度毛竹林(P<0.05)的研究结果, 说明为达到多花黄精良好生长和高效产出, 在毛竹林中进行多花黄精复合经营需要建立合理的立竹密度。 在试验毛竹林立地条件和经营水平下, 毛竹-多花黄精复合经营宜选择1 500~2 500 株·hm-2的立竹密度。 对试验林地土壤化学性质分析结果也表明随着立竹密度增加, 表层土壤中的有机质、 全氮、全磷、 全钾、 碱解氮、 有效磷、 速效钾、 pH 值均呈现出递减趋势, 以1 500~2 500 株·hm-2的立竹密度毛竹林的土壤各种营养元素含量较高。 当然, 毛竹-多花黄精复合经营, 不但要考虑多花黄精各项生产指标和经济指标, 也要考虑毛竹的各项指标, 立竹密度为1 500~2 500 株·hm-2的复合经营模式能够使多花黄精的产出效益最大化, 但对于毛竹生产的经济效益未必最大, 同时鉴于相同立竹密度条件下立竹胸径大小对林分郁闭度引起的林内光照的影响, 需进一步开展以毛竹材用林(毛竹笋用林、 笋材两用林经营强度较高, 不适合复合经营)丰产林分结构的立竹胸径条件下的毛竹-多花黄精复合经营立竹密度优化试验研究, 或开展光照对多花黄精生长和生理生态的影响多梯度控制试验, 摸清多花黄精良好生长的光照条件, 进而根据毛竹林立竹密度与林内光照强度的关系, 构建出毛竹-多花黄精复合经营的优化立竹密度。
[1] 郑郁善, 王舒凤. 杉木毛竹混交林的毛竹地下鞭根结构特征研究[J]. 林业科学, 2000, 36 (6): 69-72.ZHENG Yushan, WANG Shufeng. Study on bamboo underground structure of mixed forest of Chinese fir and bamboo[J]. Sci Silv Sin, 2000, 36 (6): 69-72.
[2] 郑郁善, 陈礼光, 洪伟. 毛竹杉木混交林生产力和土壤性状研究[J]. 林业科学, 1998, 3 (增刊): 16-24.ZHENG Yushan, CHEN Liguang, HONG Wei. Study on productivity and soil properties of mixed forests of Chinese fir and Phyllostachys pubescens [J]. Sci Silv Sin, 1998, 3 (supp): 16 - 24.
[3] 何东进, 洪伟, 吴承祯. 毛竹杉木混交林土壤团粒结构的分形特征研究[J]. 热带亚热带植物学报, 2002, 10 (3):215-221.HE Dongjin, HONG Wei, WU Chengzhen. Fractal features of soil aggregate structure under mixed plantations of Phyllostachys edulis and Cunninghamia lanceolata [J]. J Trop & Subtrop Bot, 2002, 10 (3): 215-221.
[4] 吴柏林, 陈双林, 虞敏之, 等. 毛竹纯林与竹杉混交林竹材形态质量的比较研究[J]. 浙江林业科技, 2007, 27(4): 47-50.WU Bolin, CHEN Shuanglin, YU Minzhi, et al. Comparison study on timber quality of pure bamboo forest and mixed forest [J]. J Zhejiang For Sci Technol, 2007, 27 (4): 47-50.
[5] 游秀花. 杉木毛竹混交林土壤团聚体对有机质含量的影响分析[J]. 江西农业大学学报, 2004, 26 (4): 536-539.YOU Xiuhua. A study on the effect of soil aggregate on organic matter in mixed forests of Chinese fir and Phyllostachys pubescens [J]. Acta Agric Univ Jiangxi, 2004, 26 (4): 536-539.
[6] 叶永恩. 毛竹-马尾松-杉木混交林群落学特征研究[J]. 江西农业大学学报, 2003, 25 (4): 594-598.YE Yongen. Phytocoenological features of the Phyllostachys pubescens-Pinus massoniana-Cunninghamia lanceolata community [J]. Acta Agric Univ Jiangxi, 2003, 25 (4): 594-598.
[7] 王守吉. 人工促进马尾松毛竹混交林混交效益分析[J]. 林业科技开发, 2003, 17 (增刊): 25-26.WANG Shouji. Artificial mixed-benefit analysis of the Phyllostachys pubescens and Pinus massoniana [J]. China For Sci Technol, 2003, 17 (supp): 25-26.
[8] 曹永慧, 萧江华, 陈双林, 等. 竹阔混交林阔叶树下土壤养分对毛竹生长的影响[J]. 南京林业大学学报: 自然科学版, 2007, 31 (6): 43-47.CAO Yonghui, XIAO Jianghua, CHEN Shuanglin, et al. The effect of soil nutrient around broad-leaved trees on Phyllostachys edulis growth in the mixed forest [J]. J Nanjing For Univ Nat Sci Ed, 2007, 31 (6): 43-47.
[9] 林振清. 竹阔混交林毛竹生产力与经营效益的研究[J]. 竹子研究汇刊, 2000, 19 (4): 42-45.LIN Zhenqing. Studies on bamboo productivity and advantage of mixed forest of bamboo with broad-leaved tree [J]. J Bamboo Rese, 2000, 19 (4): 42-45.
[10] 毕研文, 杨永恒, 宫俊华, 等. 黄精和多花黄精中多糖及薯蓣皂苷元的含量测定[J]. 长春中医药大学学报, 2010,26 (5): 649-650.BI Yanwen, YANG Yongheng, GONG Junhua, et al. Polysaccharide and diosgenin determination of Polygonatum sibiricum and Polygonatum cyrtonema [J]. J Changchun Univ Trad Chin Med, 2010, 26 (5): 649-650.
[11] 余红, 张小平, 邓明强, 等. 多花黄精挥发油GC2MS 分析及其生物活性研究[J]. 中国实验方剂学杂志, 2008, 14(5): 4-6.YU Hong, ZHANG Xiaoping, DENG Mingqiang, et al. Study on constituents and biological activity of volatile oil from tubers of Polygonatum cyrtonema Hua [J]. Chin J Exp Trad Med Formul, 2008, 14 (5): 4-6.
[12] 徐红梅, 赵东利. 植物生长调节剂对多花黄精芽体外发生过程中性状的影响[J]. 中草药, 2003, 34 (9): 855-858.XU Hongmei, ZHAO Dongli. Effect of plant growth regulators on several characteristics during in vitro bud regeneration of Polygonatum cyrtonema [J]. Chin Trad Herb Drug, 2003, 34 (9): 855-858.
[13] 李金花, 周守标, 王影, 等. 多花黄精5 个居群叶片的比较解剖学研究[J]. 广西植物, 2007, 27 (6): 826-831.LI Jinhua, ZHOU Shoubiao, WANG Ying, et al. Leaf comparative anatomy of Polygonatum cyrtonema from five populations [J] Guihaia, 2007, 27 (6): 826-831.
[14] 周守标, 李金花, 罗琦, 等. 多花黄精叶表皮的发育[J]. 西北植物学报, 2006, 26 (3): 551-557.ZHOU Shoubiao, LI Jinhua, LUO Qi, et al.Leaf epidermis development of Polygonatum cyrtonema [J]. Acta Bot Boreali-Occideatalia Sin, 2006, 26 (3): 551-557.
[15] 孟宪字. 测树学[M]. 北京: 中国林业出版社, 1996: 45-64.
[16] 鲁如坤. 土壤农业化学分析方法[M]. 北京: 中国农业科技出版社, 2000: 146-336.
[17] 屈卫群, 王绍华, 陈兵林, 等. 棉花主茎叶SPAD 值与氮素营养诊断研究[J]. 作物学报, 2007, 33 (6): 1010- 1017.QU Weiqun, WANG Shaohua, CHEN Binglin, et al. SPAD value of cotton leaves on main stem and nitrogen diagnosis for cotton growth [J]. Acta Agron Sin, 2007, 33 (6): 1010-1017.
[18] 钟全林, 程栋梁, 胡松竹, 等. 刨花楠和华东润楠叶绿素含量分异特征及与净光合速率的关系[J]. 应用生态学报, 2009, 20 (2): 271 - 276.ZHONG Quanlin, CHENG Dongliang, HU Songzhu, et al.Chlorophyll content and net photosynthetic rate of Machilus pauhoi and M. leptophylla [J]. Chin J Appl Ecol, 2009, 20 (2): 271 - 276.
[19] 姜丽芬, 石福臣, 王化田, 等. 叶绿素计SPAD-502 在林业上应用[J]. 生态学杂志, 2005, 24 (12): 1543-1548.JIANG Lifen, SHI Fucheng, WANG Huatian, et al. Application tryout of chlorophyll meter SPAD-502 in forest[J].Chin J Ecol, 2005, 24 (12): 1543-1548.
[20] 翟明普. 北京西山地区油松、 元宝枫混交林生物量和营养元素循环的研究[J]. 北京林学院学报, 1982, 4 (4):67-79.ZHAI Mingpu. Biomass and nutrition elements cycling studies in some mixed stands of pine and maple in Xishan area, Beijing [J]. J Beijing For Coll, 1982, 4 (4): 67-79.
[21] 毕研文, 宫俊华, 杨永恒. 光照强度对泰山黄精生长特性及产量的影响[J]. 中国农学通报, 2008, 24 (9): 315-319.BI Yanwen, GONG Junhua, YANG Yongheng. The influence of light intensity to Taishan sealwort’s growth characteristic and yield [J]. Chin Agric Sci Bull, 2008, 24 (9): 315-319.
[22] 樊后保, 李燕燕, 苏兵强, 等. 马尾松-阔叶树混交异龄林生物量与生产力分配格局[J]. 生态学报, 2006, 26 (8):2463-2473.FAN Houbao, LI Yanyan, SU Bingqiang, et al. Allocation pattern of biomass and productivity in the mixed unevenaged stands of masson’s pine and hardwood species[J]. Acta Ecol Sin, 2006, 26 (8): 2463-2473.
猜你喜欢
天津农林科技(2022年2期)2022-04-19
中老年保健(2021年11期)2021-08-22
浙江农林大学学报(2020年5期)2020-11-05
Digital Chinese Medicine(2020年3期)2020-11-03
云南农业科技(2020年5期)2020-10-14
意林·少年版(2020年13期)2020-08-02
汉语世界(2020年3期)2020-06-19
东坡赤壁诗词(2020年2期)2020-06-04
湖州职业技术学院学报(2020年4期)2020-03-30
饮食科学(2009年3期)2009-03-23
