
TLR7、TLR8在非小细胞肺癌中的表达及其临床意义!
2013-05-16 03:11马海丽逯清丽
华中科技大学学报(医学版) 2013年3期
马海丽, 刘 建, 逯清丽
华中科技大学同济医学院附属同济医院综合科,武汉 430030
肺癌是导致人类死亡的主要疾病之一,虽然近年来在肺癌的临床治疗方面有很大的进展,但肺癌的5年生存率仍相对较低,为5%。Toll样受体(Toll like receptors,TLRs)最早发现于果蝇体内,是一类模式识别受体,识别病原和损伤相关分子模式,在启动天然免疫应答的同时,也参与激活特异性免疫应答,在宿主免疫防御中起着重要作用。近来研究表明,TLRs在肿瘤形成、免疫治疗方面起着一定的作用。TLR7、TLR8作为TLRs的成员,主要识别单链RNA,参与病毒感染[1-2]。对其和肺癌的关系目前研究尚少。本研究采用免疫组化方法,检测TLR7、TLR8在非小细胞肺癌中的表达,分析其与肺癌临床病理特征之间的关系,初步探讨TLR7、TLR8在非小细胞肺癌中的作用,为进一步研究TLR7、TLR8在肺癌中的治疗应用提供理论依据。
1 资料与方法
1.1 临床资料
随机选取2010~2011年华中科技大学同济医学院病理学系保存的43例非小细胞肺癌及其中20例的癌旁正常肺组织标本(距离肿瘤边缘5cm以上)。所有患者在手术之前均未接收放疗或化疗,排除慢性阻塞性肺疾病、肺炎等呼吸系统疾病,并且具有完整的病理资料。所有标本均经病理检查证实为非小细胞肺癌或癌旁正常肺组织。患者平均年龄为54.4岁,男性患者34例,女性患者9例。其中鳞癌21例,腺癌22例;分化程度为高中分化27例,低分化16例;依据国际抗癌联盟UICC肺癌TNM分期(2009年版)分为Ⅰ期12例,Ⅱ期8例,Ⅲ期22例,Ⅳ期1例;淋巴结转移者23例,无淋巴结转移者20例。
1.2 主要试剂
兔抗人TLR7单克隆抗体、小鼠抗人TLR8多克隆抗体均购于启动子生物有限公司。过氧化物酶标记的链霉卵白素(SP)通用型试剂盒、DAB显色试剂盒购于北京中杉金桥生物有限公司。苏木精染液购于武汉谷歌生物有限公司。
1.3 实验方法
采用免疫组化SP三步法。所有蜡块均连续4 μm切片,切片置于60℃恒温烤箱中20min,脱蜡水化。PBS冲洗后,3%H2O2室温孵育30min,PBS冲洗,进行微波抗原修复20min,PBS冲洗后,正常山羊血清室温封闭20min左右。加入兔抗人TLR7抗体(1∶50稀释),鼠抗人TLR8抗体(1∶100稀释)一抗后,置于4℃冰箱中孵育过夜。次日取出复温45min后,加入生物素标记的二抗和辣根酶标记的链霉卵白素工作液,DAB显色后,苏木精复染,脱水,透明,中性树胶封片。PBS缓冲液代替一抗作为阴性对照。
1.4 结果判定
根据文献[3]标准,TLR7、TLR8以细胞质中出现棕黄色或棕褐色着色为阳性,400倍光镜下计数,免疫组化结果依阳性细胞数和染色强度进行综合判定。阳性细胞数计分,依阳性细胞率≤10%、11%~、26%~、51%~、>75%分别记为0~4分;染色强度记分,依无着色、浅棕黄色、棕黄色、棕褐色分别记为0~3分。二者得分相乘,大于等于平均分者为阳性表达,小于者判为阴性表达。
1.5 统计学分析
应用SPSS 12.0统计分析软件,对实验结果进行卡方检验及Pearson相关性分析,以P<0.05为差异具有统计学意义。
2 结果
2.1 TLR7、TLR8在非小细胞肺癌组织及正常肺组织中的表达
TLR7、TLR8的阳性表达主要以细胞质中出现棕黄色或棕褐色着色为标准。TLR7主要表达在肺癌细胞核周围(图1),在非小细胞肺癌组织中的阳性表达为48.8%(21/43),高于癌旁正常肺组织(3/20,15.0%),差异具有统计学意义 (P<0.05)。TLR8表达在肺癌细胞的细胞质中(图2),相对TLR7,表达分布弥散,在非小细胞肺癌中的阳性表达为69.8%(30/43),高于癌旁正常肺组织(4/20,20.0%),差异具有统计学意义(P<0.05)。正常肺组织中,TLR7和TLR8在正常肺泡上皮细胞和肺泡巨噬细胞中有少量表达(图3)。
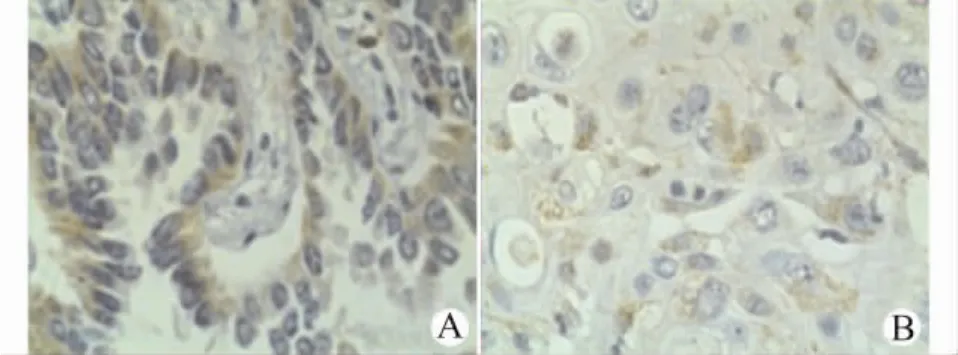
图1 TLR7在肺腺癌(A)和肺鳞癌(B)中的表达(DAB显色,×400)Fig.1 The expression of TLR7in lung adenocarcinoma(A)and squamous cell carcinoma(B)(DAB,×400)

图2 TLR8在肺腺癌(A)和肺鳞癌(B)中的表达(DAB显色,×400)Fig.2 The expression of TLR8in lung adenocarcinoma(A)and squamous cell carcinoma(B) (DAB,×400)

图3 TLR7(A)、TLR8(B)在正常肺组织中的表达(DAB显色,×400)Fig.3 The expression of TLR7(A)and TLR8(B)in the normal lung tissue(DAB,×400)
2.2 TLR7、TLR8表达与非小细胞肺癌临床病理特征之间的关系
TLR7的表达与性别和病理类型有关,与吸烟、年龄、淋巴结转移、TNM分期、分化程度无相关性。TLR8的表达与吸烟有关,但与年龄、性别、病理类型、淋巴结转移、TNM分期、分化程度等无相关性(表1)。
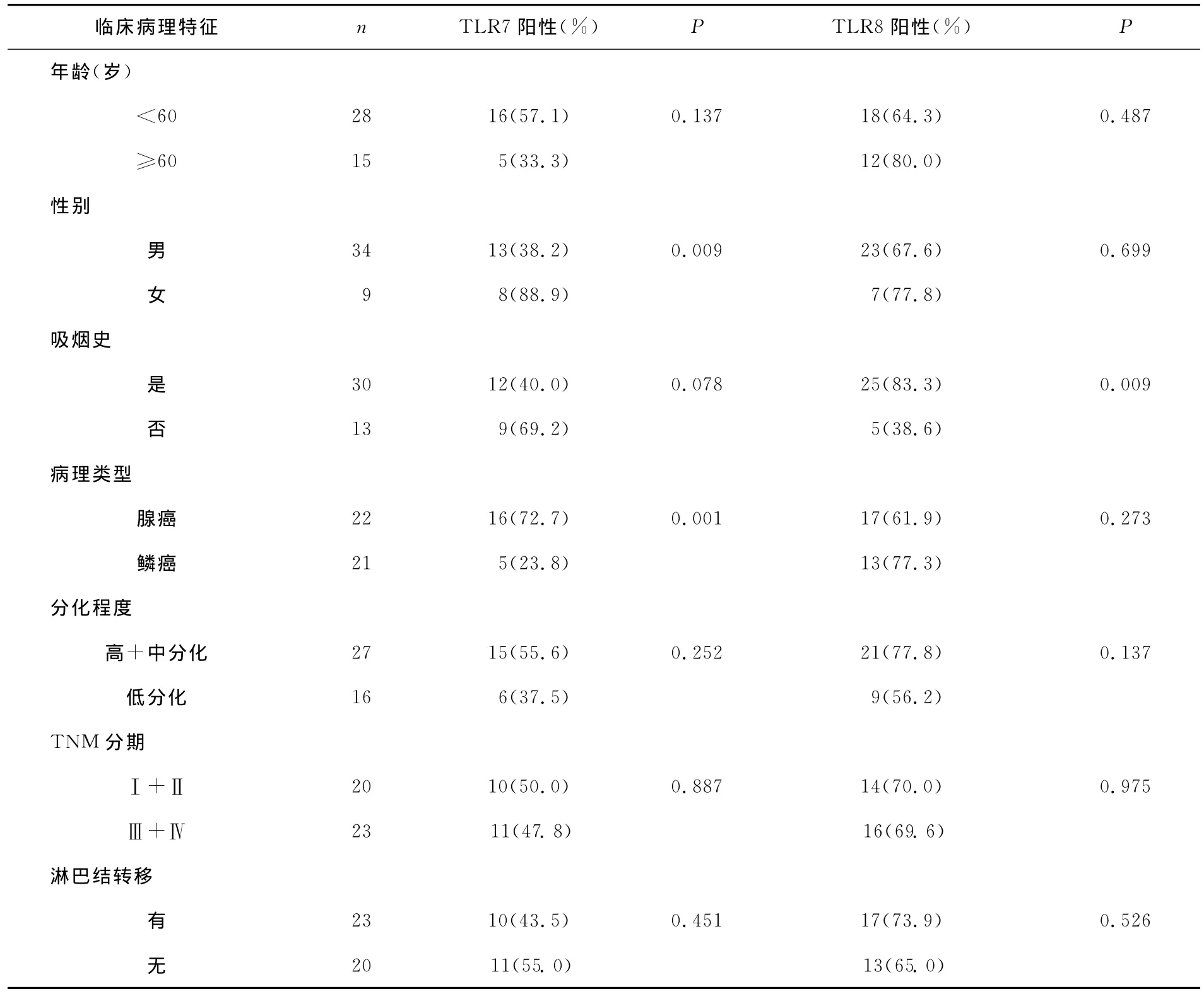
表1 TLR7、TLR8与非小细胞肺癌临床病理特征之间的关系Table 1 The relationship between expressions of TLR7and TLR8and clinicopathologic features of NSCLC tissues
2.3 非小细胞肺癌中TLR7和TLR8表达的相关性
TLR7和TLR8在43例非小细胞肺癌组织中的表达,均阳性的16例,均阴性的8例,TLR8阳性而TLR7阴性的14例,TLR8阴性而TLR7阳性的5例,Pearson相关分析结果表明,TLR7、TLR8在肺癌中的表达无相关性(P=0.382)。
3 讨论
Toll基因最早发现于果蝇体内,编码Toll蛋白。人们发现Toll及Toll信号传导通路有助于果蝇对抗感染过程,其主要作为受体抵抗真菌感染[4]。后来人们相继在植物、动物、人类体内发现了类似果蝇体内Toll的受体及其信号通路,因此,此类受体被称为 Toll-like receptors(TLRs)。目前在人类体内至少发现10个TLR家族成员。TLRs胞外富含亮氨酸重复片段,以识别配体,各个TLR家族成员识别的病原成分不同。TLRs识别病原(体外)和损伤(体内)相关分子模式,前者包括细菌、病毒、真菌等病原,后者主要是体内炎症、损伤信号。TLRs信号途径释放NF-κB、IFN-α等一系列因子,上调抗原递呈细胞表面的共刺激分子表达,启动天然免疫应答,激活非特异性免疫应答[5-7],参与机体免疫防御过程。TLR7、TLR8作为TLRs家族成员,两者具有相似的结构[8],其主要定位表达于细胞质内,TLR7主要表达于抗原提呈细胞(包括CD34+树突状细胞),TLR8主要表达于单核细胞、巨噬细胞、及髓系树突状细胞[9]。其配体包括天然配体和人工合成的化合物两种。前者是病毒复制过程中产生的单链RNA,介导病毒感染过程;后者包括人工合成的RNA片段及咪唑喹啉胺类等。咪喹莫特作为一类重要的咪唑喹啉胺类成员,已应用于临床病毒感染及某些皮肤肿瘤的局部免疫治疗[10]。
有研究表明,TLRs不仅在免疫细胞表达,其在肿瘤细胞上也存在表达。TLR信号传导通路启动NF-κB、MAPK信号通路,释放炎症因子,在炎症、肿瘤的形成、发展过程中起着重要作用。有研究发现,TLRs表达的增加,一定程度上和食管癌等肿瘤的发生、发展有关[3,11]。Ochi等[12]发现 TLR7在人和小鼠的胰腺癌组织中表达增加,经其配体刺激后,上调p53、c-Myc、TGF-β等基因的表达,明显加速肿瘤进展。Cherfils-Vicini等[13]用相应配体刺激肺癌细胞株上的TLR7和TLR8后,观察到NF-κB激活,抗凋亡蛋白Bcl-2蛋白表达上调。TLR家族成员TLR4和TLR9也被发现在肺癌组织中存在高表达[14]。同时,有研究发现相对于正常肺组织,在非小细胞肺癌细胞中TLR7mRNA水平明显增加[15]。这些研究提示TLRs可能与肿瘤的发生、发展密切相关。
目前对于TLR7和TLR8在肺癌组织中的表达研究尚少。本研究采用免疫组化方法检测了TLR7和TLR8在非小细胞肺癌及癌旁正常组织中的蛋白质表达水平,结果显示TLR7在鳞癌及腺癌中的阳性表达率为48.8%,TLR8阳性表达率为69.8% ,两者均高于癌旁正常肺组织,这与Cherfils-Vicini等[13]的结果是一致的。本研究显示,TLR7在腺癌组织中的阳性表达率为72.7%,高于鳞癌组织中的阳性表达率23.8%,两者之间差异具有统计学意义,表明TLR7在非小细胞肺癌的表达与病理类型有关。同时发现女性肺癌患者的TLR7表达阳性率高于男性肺癌患者,这可能与腺癌女性多发有关。有吸烟史的肺癌患者癌组织中TLR8阳性表达率高于无吸烟史者,提示吸烟可能是影响TLR8参与肺癌进程的一个危险因素。本研究未发现TLR7、TLR8的表达与肺癌分化程度、病理分期、淋巴结转移之间的相关性,可能与样本例数较少有关,因此需进行更大样本量的研究。在TLRs家族中,TLR7和TLR8有着相似的结构,本研究未发现两者之间有相关性,推测两者可能在肺癌发展过程中没有协同作用。
总之,TLRs与其相应配体结合后,释放一系列的炎症、细胞因子,参与肿瘤形成、发展过程,TLR7和TLR8在非小细胞肺癌细胞中高表达,提示两者参与肺癌发病过程,其具体机制及信号通路尚需进一步研究。
[1] Thompson M R,Kaminski J J,Kurt-Jones E A,et al.Pattern recognition receptors and the innate immune response to viral infection[J].Viruses,2011,3(6):920-940.
[2] Lund J M,Alexopoulou L,Sato A,et al.Recognition of singlestranded RNA viruses by Toll-like receptor 7[J].Proc Natl Acad Sci USA,2004,101(15):5598-5603.
[3] Sheyhidin I,Nabi G,Hasim A,et al.Overexpression of TLR3,TLR4,TLR7and TLR9in esophageal squamous cell carcinoma[J].World J Gastroenterol,2011,17(32):3745-3751.
[4] Lemaitre B,Nicolas E,Michaut L,et al.The dorsoventral regulatory gene cassette spatzle/Toll/cactus controls the potent antifungal response in Drosophila adults[J].Cell,1996,86(6):973-983.
[5] Pasare C,Medzhitov R.Toll-like receptors:linking innate and adaptive immunity[J].Microbes Infect,2004,6(15):1382-1387.
[6] 陈华文,祝伟,李树生.内毒素预处理对内毒素血症大鼠肝TLR-4、NF-κB和ICAM-1表达的影响[J].华中科技大学学报:医学版,2011,40(6):666-669,685.
[7] 王家顺,张俊,王建军,等.TOLL样受体2在大鼠肺移植缺血再灌注损伤期的时序性表达及意义[J].华中科技大学学报:医学版,2011,40(2):165-168.
[8] Roach J C,Glusman G,Rowen L,et al.The evolution of vertebrate Toll-like receptors[J].Proc Natl Acad Sci USA,2005,102(27):9577-9582.
[9] 杨恒.TLR7/8激动剂及其在肿瘤免疫治疗中的研究进展[J].肿瘤防治研究,2011,38(4):471-473.
[10] Adams S.Toll-like receptor agonists in cancer therapy[J].Immunotherapy,2009,1(6):949-964.
[11] Coussens L M,Werb Z.Inflammation and cancer[J].Nature,2002,420(6917):860-867.
[12] Ochi A,Graffeo C S,Zambirinis C P,et al.Toll-like receptor 7 regulates pancreatic carcinogenesis in mice and humans[J].J Clin Invest,2012,122(11):4118-4129.
[13] Cherfils-Vicini J,Platonova S,Gillard M,et al.Triggering of TLR7and TLR8expressed by human lung cancer cells induces cell survival and chemoresistance[J].J Clin Invest,2010,120(4):1285-1297.
[14] Zhang Y B,He F L,Fang M,et al.Increased expression of Toll-like receptors 4and 9in human lung cancer[J].Mol Biol Rep,2009,36(6):1475-1481.
[15] Samara K D,Antoniou K M,Karagiannis K,et al.Expression profiles of Toll-like receptors in non-small cell lung cancer and idiopathic pulmonary fibrosis[J].Int J Oncol,2012,40(5):1397-1404.
猜你喜欢
云南医药(2021年3期)2021-07-21
四川文理学院学报(2020年5期)2020-02-12
中成药(2018年7期)2018-08-04
材料科学与工程学报(2016年4期)2017-01-15
西南军医(2016年3期)2016-01-23
合成化学(2015年4期)2016-01-17
中国当代医药(2015年17期)2015-03-01
癌变·畸变·突变(2015年3期)2015-02-27
西南军医(2014年5期)2014-04-25
无机化学学报(2014年6期)2014-02-28
