
云锦杜鹃成花逆转研究
2013-05-11 07:19张云生边才苗
浙江林业科技 2013年4期
张云生,边才苗
(1. 浙江省台州市椒江区林业特产总站,浙江 台州 318000;2. 台州学院 生态研究所,浙江 台州 318000)
云锦杜鹃成花逆转研究
张云生1,边才苗2
(1. 浙江省台州市椒江区林业特产总站,浙江 台州 318000;2. 台州学院 生态研究所,浙江 台州 318000)
成花逆转是植物从生殖生长状态逆向转变为营养生长,是植物生长发育过程中的特殊现象。云锦杜鹃(Rhododendron fortunei),其花芽发育早期有高比例的成花逆转,主要是花序逆转,即在花器官分化前,花芽的发育中断并提前爆芽,形成只有叶片的营养短枝,少量花逆转。研究结果表明:幼树的爆芽率很高,尤其是低海拔地区栽种,爆芽率为16.82%,显著高于自然条件的10.42%;成株的爆芽率较低,自然条件下只有4.17%,单纯的去幼果可以提高花芽的发生率,但不增加爆芽率,结合施肥,爆芽率也显著增加,达到 9.09%,说明个体的年龄、栽培的气候条件和个体的营养状况是诱发爆芽的主要原因,其中营养物质很可能是过量积累。
云锦杜鹃;花芽;花序逆转;爆芽率
云锦杜鹃(Rhododendron fortunei)树高2 ~7 m,为常绿灌木或小乔木[1],花期5月,花大如碗,花冠艳丽,且有“千花杜鹃”之美称,是一种观赏价值高、开发潜力大的园林树种。云锦杜鹃原产我国,分布于长江流域各省,大多零星地散落在海拔400 ~ 1 000 m的沟谷阔叶林中或山顶灌草丛。浙江省天台县华顶国家森林公园有20 hm2连片的云锦杜鹃群落,有2万多株,其中树龄500 a以上的108株,堪称华东一绝。近年来,这片杜鹃林呈现生长势减弱、开花指数降低等衰退现象[2~3]。为此,园林部门采取了以下措施:秋季清理阔叶箬竹等灌草层植物,开春前后2次根区深施复合肥,并在花后摘除幼果,以改善花芽发育的资源供应。通过整治,云锦杜鹃的开花状况有了明显的改观,但花芽的提前爆芽现象也显著增加,即部分“花芽”的发育受阻,演变为枝芽在秋季提前展开,形成营养短枝。基于这种提前爆芽属于花序逆转(inflorescence reversion)的后续表现;同时,云锦杜鹃的花芽发育期很长,有利于从代谢及超微结构方面确定成花转变的细节变化。本文观察和统计了云锦杜鹃的花序逆转现象,并分析了诱发这种逆转的主要原因,以期为后续的深化研究提供基础依据和信息。
1 材料与方法
1.1 材料
选择云锦杜鹃2个年龄段的个体:①幼树的树龄为10 ~ 12 a,开花龄2 a以上,其中栽培个体取材于浙江省台州学院,28° 45′ N,120° 31′ E,海拔40 m,种植在大棚内(用单层黑色纱网遮盖,透光度70%,同时大气相对湿度在 75%左右);自然种群位于浙江省天台县华顶国家森林公园,29° 15′ N,121° 06′ E,主峰海拔1 098 m,样地在主峰西侧的观云亭附近,海拔850 m;②成株的树龄为30 ~ 40 a,其中自然条件和去幼果处理在主峰北侧黄经洞,海拔900 m;去幼果+施肥处理在主峰西侧的永字亭附近,海拔820 m。各样地的主要环境资料参见文献[4~5]。
1.2 方法
1.2.1 试验处理 于2011年11月在每个样地内选择5株样树标记,施肥为2次(12月和次年3月),所施肥料为复合肥(15-15-15);去幼果处理在5月下旬进行。由于永字亭周围是华顶国家森林公园的观赏区,该样地的去幼果和施肥是园林工人完成的。
1.2.2 花芽发育进程观察 根据云锦杜鹃花芽分化发育的规律,于6月初花期结束后开始,每月1次测量标记样树上的顶芽大小(直径和高度),并将其作为圆锥体估算其体积;在9月中旬花芽基本成形后,统计花芽数、当年生营养枝和花枝的数量,其中幼树为整株,成株为东南西北四个方位各1个4级枝条;提前爆芽的数量统计时间为10月10日前后,同时摄影记录爆芽发生时的形态,并取非标记枝上的正常花芽,石蜡切片观察花芽的发育期。
1.2.3 数据计算 云锦杜鹃的花芽主要来自顶芽,花芽的发生率和形成率计算均以顶芽为总体,其中发生率是9月中旬的统计结果,而形成率是爆芽发生后的统计结果;同时,爆芽率只统计来自顶芽的花芽的爆芽数量,相应的计算公式如下:

2 结果与分析
2.1 花芽的发生与发育
云锦杜鹃性喜光,在天台山,主要分布于海拔800 ~ 1 000 m的阳坡和半阴坡。开春后,枝芽迅速膨大,4月下旬开始展露枝叶,随后花芽也开始活动,4月底显露花蕾,5月初开花,花期近1个月;到花期结束时,当年生枝条成为长8.2 ~ 12.8 cm、有8 ~ 12叶片的营养枝,其中,多数枝条只有顶芽活动,少数枝条(约35%)还有近顶端2 ~ 5个侧芽的活动。活动芽都包含有数层芽鳞(图1-2),其中顶芽可以发育为花芽,而侧芽一般为枝芽,只有个别侧芽发育成花芽,形成1枝2个花芽(图1-1);在幼树阶段,13.7%的花芽来自于侧芽,而成年树只有7.5%。

图1 云锦杜鹃的成花逆转现象Figure 1 The flowering reversion of Rh. fortunei
云锦杜鹃花芽的发育期很长,其活动从花期结束(6月初)就开始,7-9月为旺盛生长期,芽体的体积倍增,10月为花器官的形态分化期,生长减缓,至11月生长基本停止,经过5个多月的生长和分化,芽体的体积增大了35倍,形成直径1.4~1.8 cm,高2.8 ~ 3.3 cm,内含8 ~ 12朵小花的花芽越冬(图 2)。去幼果和施肥处理的花芽略为大一些,但与对照差异很小,7个测定时间测定值的最大差异只有8.5%。而枝芽的生长不甚明显,到越冬前,芽体的直径只有0.8 cm,高度不超过1.4 cm,体积仅增加3倍。
2.2 成花逆转及其表现
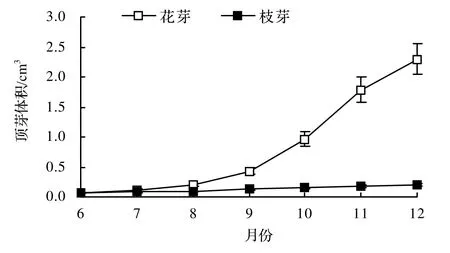
图2 云锦杜鹃顶芽大小的月变化Figure 2 Monthly changes of terminal bud’s size in Rh. fortunei
云锦杜鹃花芽的提前爆芽发生于10月上旬,此时,正常花芽的直径为1.1 ~ 1.4 cm,高2.1 ~ 2.4 cm;其花原基已开始花器官的分化,雄蕊有花药形态,小孢子母细胞也开始形成,有的已完成减数分裂(图1-9);雌蕊也有明显的子房、花柱和柱头区分(图1-8)。基于爆芽后形成的枝条是没有花芽的营养短枝(图1-3和图1-4),长度还不到5 cm,与正常营养枝区别明显;且短枝基部有明显不同于正常叶片的苞叶(图1-3),这可能是花器官发育的遗迹。因此,云锦杜鹃的提前爆芽现象属于花序逆转的后续表现,花序逆转的发生期可能在9月下旬或更早;同时,逆转前的分化活动对其原有的营养生长属性也有很大的影响。
另外,云锦杜鹃还有少量的花逆转,这种逆转芽没有提前爆芽的表现,在花期爆芽后形成的枝条为上部带叶,下部为正常的花,并可以正常结实(图1-5);但这种枝条相对较长,一般为6 ~ 8 cm,且下部叶片的形态或叶脉有明显的异常(图1-6和图1-7)。说明花逆转并不是轻度的成花逆转,只是这种逆转变化主要集中在芽体上部。
2.3 花芽发育及其逆转的原因
云锦杜鹃花芽发生率和爆芽率的统计结果见表1。由表1知,这2个指标主要取决于以下几个方面的因素:①个体的年龄,在自然状态下,幼树阶段的数值均显著高于成树阶段;②气候因素,在低海拔的栽培条件,2个指标值比自然条件的相应值要高一些,其中爆芽率的差异为显著水平,但花芽发生率差异不显著,因而花芽的净形成率差异很小,自然条件为57.85%,略高于栽培条件的56.88%;③个体的营养状况,单纯去幼果可以提高花芽的发生率,爆芽率也有一定的增加,但差异不明显;若结合施肥,花芽发生率和爆芽率均显著增加,但花芽形成率为52.23%,与单纯去果处理(51.24%)相差很小。

表1 云锦杜鹃花芽的发生率和爆芽率Table 1 The occurrence rate and exploded rate of floral bud in Rh. fortunei%
3 讨论
3.1 云锦杜鹃的成花决定与成花逆转
成花决定是指感受态植株接受外界成花信息后,生长点内部发生一系列的变化,部分细胞以后将转向生殖生长的一种状态[6]。引起成花决定的条件包括植物体内的启动因素,如植株年龄或营养生长的节数;以及体外条件,如特定的日长和温度[7]。本研究显示,云锦杜鹃的成花决定主要取决于个体的年龄,树龄30 ~ 40 a的个体花芽发生率显著下降,这可能与个体生殖力的减退有关;同时,改善植株的营养状况(去幼果和施肥)均会显著提高花芽的发生率,说明资源限制是影响其花芽发生的重要因素。另外,在低海拔地区栽种的幼树,花芽的发生率和形成率并不比高海拔的自然条件低,说明夜低温和日长(华顶山因云雾弥漫,日长缩短2 h左右)等气候因子不是云锦杜鹃成花转变的决定要素。
一般而言,植物完成成花转变后,顶端分生组织开始稳定的生殖生长,但如果环境条件不适合,植物也可能从生殖生长状态逆向转回营养生长,这种现象称为成花逆转(flowering reversion)。成花逆转有3种类型[8]:花逆转(floral reversion)、花序逆转(inflorescence reversion)和部分开花(partial flowering)。云锦杜鹃主要是花序逆转,并于10月上旬提前爆芽,形成没有花芽的带叶营养短枝,同时还有少量的花逆转。基于花逆转在花期爆芽,形成的枝条下部为正常的花,但上部为带叶的营养茎,相对较长,且下部的叶片有异常形态。提示花逆转的发生可能更早,且芽体顶端的逆转可以阻止这种异常的继续延伸。
3.2 云锦杜鹃成花逆转的原因分析
成花逆转与环境条件有关,是植物对环境条件的一种应答。不同种类的植物,逆转由不同的因素引起,有时由多个因素引起。一般来说,引起逆转的环境因素与诱导条件相反,在短日植物大豆的一些晚熟品种,如自贡冬豆[9]和野大豆[10]经短日处理诱导开花,但开花后若置于长日照下,生殖生长将受到明显的抑制,恢复旺盛的营养生长状态;典型的亚热带果树龙眼,不仅花序的生理分化期需要低温,花序发育过程也同样需要一段低温期,才能使花序发育正常[11]。表明植物的成花转变是一个量变过程,而不是质变过程[12],或者说,成花诱导的前期效应具有可逆性[9]。
根据营养物质转移学说(nutrient diversion hypothesis),诱导处理使同化物向顶端分生组织积累并导致开花[13]。本研究显示,改善植株的营养状况,花芽的发生率显著增加,但提前爆芽率也有一定的提高,尤其是去幼果+施肥处理;且多数逆转芽是没有活动侧芽的单生顶芽。基于花器官分化前,芽体中蔗糖和葡萄糖的含量是枝芽最高,单生顶端花芽其次,有活动侧芽的顶端花芽最低(数据将另文报道)。说明顶芽中同化物积累过多同样也不利于花芽的发育,甚至还可能导致发育的逆转。
诱导植物成花的原因是十分复杂的,多因子控制模型(multifactorial control model)认为,营养物质的积累只是诱导开花的一个因素,另外还有大量的诱导物和抑制物在开花过程中起作用;即只有限制与诱导因子在适宜时期达到一定浓度,形成新的平衡后才会开花[13]。基于云锦杜鹃成花逆转后形成的枝条为短枝,提示其成花转变过程可能有大量的生长抑制物质产生,但这些尚有待于试验证实。另外,植物是否已完成成花决定需要观察其后效,目前尚未能从代谢及超微结构观察中确定成花决定的指标[14]。云锦杜鹃的花芽发育期很长,有利于捕捉到花芽发育过程的异常形态和生理变化。
[1] 丁炳扬,方云亿. 浙江杜鹃花属植物的研究[J]. 杭州大学学报,1989,16(2):194-200.
[2] 管康林,吴家森,范义荣,等. 华顶山云锦杜鹃林衰退原因及对策[J]. 浙江林学院学报,2001,18(2):195-197.
[3] 吴家森,庞加钱,周祖耀,等. 华顶山云锦杜鹃林地土壤肥力分析[J]. 浙江林业科技,2002,22(2):26-28.
[4] 边才苗,金则新. 云锦杜鹃的开花动态与繁育系统研究[J]. 广西植物,2005,25(2):169-173.
[5] 边才苗,金则新. 天台山云锦杜鹃的开花与结实特性[J]. 园艺学报,2006,33(1):101-104.
[6] McDaniel C N,Siger S R,Smith S M E. Development states associated with the floral transion[J]. Dev Biol,1992(153):59-69.
[7] Bernier G,Havelange A,Houssa C,et al. Physiological signals that induce flowering[J]. Plant Cell,1993(5):1 147-1 155.
[8] Battey N H,Lyndon R F. Reversion of flowering[J]. Bot Rev,1990(56):162-189.
[9] 韩天富,王金陵. 大豆开花后光周期反应的研究[J]. 植物学报,1995,37(11):863-869.
[10] 周三,赵可夫. 耐盐野生大豆(Glycine soja)的光周期效应[J]. 植物生理与分子生物学学报,2002,28(2):145-152.
[11] 柯冠武,黄进华,王长春. 龙眼花穗“冲梢”形态解剖学的观察[J]. 中国果树,1988(1):22-24.
[12] 傅永福,孟繁静. 植物成花转变过程的基因调控[J]. 植物生理学通讯,1997,33(5):393-400.
[13] Bernier G. The control of floral evocation and morphogenesis[J]. Annu Rev Plant Physiol Plant Mol Biol,1988(39):175-219.
[14] 白素兰,谢中稳,刘永胜,等. 植物的成花逆转[J]. 植物生理学通讯,2000,36(3):252-257.
Observations on Flowering Reversion of Rhododendron fortunei
ZHANG Yun-sheng1,BIAN Cai-miao2
(1. Jiaojiang Forestry Specialty Station of Zhejiang, Taizhou 318000, China; 2. Institute of Ecology, Taizhou University, Taizhou 318000, China)
Observations were conducted on flowering reversion of young and adult Rhododendron fortunei in Tiantai, Zhejiang province. The result demonstrated high proportion of flowering reversion in the early development of floral buds of Rh. fortunei, with the inflorescence reversion being the mainly type. Statistics analysis showed the exploded buds rate of young trees, especially of that planted in the low-altitude(16.82%) was higher than that in the natural conditions (10.42%). The exploded rate of adult trees was only 4.17% under the natural condition. Removal of young fruits (unfertilizred) could significantly increase the occurrence rates of floral bud (from 48.9% to 54.6%), exploded rate was not significant. But when the adult trees were fertilized, removal of young fruits would increase exploded rate to 9.09%. The observations resulted that age, climate and nutritional status of each tree were major factors of the explosion bud. And the nutrients may be excessive accumulated.
Rhododendron fortunei; floral bud; inflorescence reversion; rate of exploded buds
S718.43
A
1001-3776(2013)04-0018-05
2013-04-10;
2013-05-25
张云生(1965-),男,浙江温岭人,高级工程师,从事森林生态研究。
猜你喜欢
华人时刊(2022年15期)2022-10-27
扬子江诗刊(2022年1期)2022-01-10
金山(2021年10期)2021-11-02
安徽农业科学(2021年11期)2021-07-16
安徽农学通报(2021年10期)2021-06-17
健康之家(2021年19期)2021-05-23
华人时刊(2020年13期)2020-09-25
农民致富之友(2020年22期)2020-09-02
热带生物学报(2019年1期)2019-04-18
现代农业科技(2016年9期)2016-10-20
