
水杨酸对烟草野火病的抑制效果及其对烟株的生物学效应
2013-03-14 08:35:41王振国肖鹏陈秋双李栋梁王飞尹朝先李石力丁伟
中国烟草学报 2013年5期
王振国,肖鹏,陈秋双,李栋梁,王飞,尹朝先,李石力,丁伟.4
1 中国烟草总公司重庆市公司奉节分公司,重庆 奉节 404600;2 西南大学植物保护学院,重庆 北碚 400716;3 奉节县农委植物保护与植物检疫站,重庆 奉节 404600;4 重庆烟草科学研究所,重庆 北碚 400716
烟草野火病(Pseudomonas syringaepv. tabaci) 是一种细菌性叶部病害,具有爆发性和破坏性,是影响烟叶生产的主要病害之一。其药剂防治研究结果普遍都表现为[1-3]:室内活性测定效果较好,大田推广使用时,由于受到药剂持效期、气候条件、病原抗药性的产生等因素的影响,导致其防治效果不够理想。当前,生产上关于野火病的防治现状为:单独使用一种或两种传统的化学杀菌剂已经很难有效控制野火病的发生和流行,且在野火病的防治过程中大量使用抗生素类药剂,此类药剂持效期长、残留性强,对环境安全、烟叶及其制品的安全性具有较大的潜在危害性。
水杨酸 (Salicylic acid, SA) 是植物体内的小分子物质,其化学名称为邻羟基苯甲酸。由于水杨酸由植物自身合成,含量较低,可在韧皮部运输,在植物生长过程中起着重要的调节作用[4-5],很多专家学者将其视为植物激素家族的一新成员[6],并且水杨酸还能诱导植物产生抗病性以及抵抗不良因素造成的伤害[7-9]。近期关于水杨酸诱导植物抵抗病害的研究很多,如王瑞霞等用平板法和气相色谱-质谱联用(GC-MC)方法研究了水杨酸诱导处理水稻后产生的抗菌物质对稻瘟病 (Magnaporthe grisea) 孢子萌发的影响,结果表明水杨酸处理后的水稻叶片提取物质对稻瘟病菌孢子萌发具有明显的抑制作用[10];谢鑫等通过用不同浓度的水杨酸处理烟草以测定其对TMV抑制的最佳浓度,发现8 mmol/L的水杨酸对TMV的抑制效果最好,且在诱导处理后7 d达到最佳为61.29%[11];杜小凤等发现不同浓度的水杨酸处理对黄瓜根结线虫都具有一定的防治效果[12]。
针对烟草生产上野火病危害严重、防治困难的现状,并在前人对水杨酸研究成果的基础之上,以寻求控制烟草野火病的最佳物质为目的,研究了水杨酸诱导烟草对野火病的抑制作用及其对烟株主要生物学性状的影响。
1 材料与方法
1.1 材料
1.1.1 供试药剂
水杨酸:分析纯,成都市科龙化工试剂厂;赤霉素:0.15%水剂,贵州省遵义泉通化工厂;胺鲜酯:1.6%水剂,郑州农达生化制品厂;农用链霉素:72%可湿性粉剂,石家庄通泰生化有限公司。
1.1.2 供试品种
贵烟4号。
1.1.3 供试地块
试验于2012年在重庆市奉节县太和乡石盘村选取具有代表性的地块进行 (海拔1310 m,北纬30°38′,东经109°12′)。地块相对比较平整,肥力中等、均匀,土壤粘性、有机质及微量元素含量中等,海拔1290 m,该地块有多年种烟历史。5月23日移栽,株距为60 cm,行距为115 cm。6月下旬进入团棵期,7月下旬进入打顶期。
1.2 方法
1.2.1 试验设计
处理1:水杨酸,2000倍液,使用前用少量酒精溶解后再兑水混用;处理2:0.15%赤霉素水剂,1000倍液;处理3:1.6%胺鲜脂水剂,1500倍液;处理4:72%农用链霉素可湿性粉剂,1200~1500倍液 (推荐剂量);处理5:清水对照。以上处理,施药方法均为叶面喷雾。
每个处理设置3个重复,每个小区45~50 m2(约70株烟),随机排列,设置保护行。处理1至处理3,以及处理5均于伸根期 (6月5日) 进行第一次处理,于团棵期 (6月28日) 和旺长期 (7月15日) 进行第二、三次处理;处理4团棵期 (6月28日) 进行第一次施药处理,旺长期 (7月15日) 进行第二次施药,共2次。施药后,保证药液均匀分布于烟叶的正反两面、以雾滴不下滴为宜。每次施药处理后6~8小时内无有效降雨,否则重新施药。试验过程中不喷施杀菌剂,其他农艺措施同当地常规操作。
1.2.2 病情调查
每个小区采用“对角线五点法”选点,每点挂牌定5株,每小区固定调查25株,共调查3次。第一次药后调查于6月26日进行,第二、三次药后调查分别于7月9日和7月25日进行。病害分级标准参照GB/T23222-2008《烟草病虫害调查与分级》中的烟草野火病部分。利用公式(1)计算病叶率,利用公式 (2)计算病情指数,利用公式 (3) 计算防治效果,用SPSS软件中的“新复极差—邓肯”法对病情指数、诱抗(防治) 效果进行方差分析。

1.2.3 生物学性状调查
按 YC/T142-1998《烟草农艺性状调查方法》对烟株的叶片数、最大腰叶长、最大腰叶宽、株高以及茎围进行调查,并利用公式 (4)、(5) 计算叶面积。于团棵期(6月27日)、打顶期(7月26日),共调查两次。调查时,采用“对角线五点法”选点,每点定5株,每个小区定株调查25株。

2 结果
2.1 水杨酸对烟草野火病的控制效果
水杨酸对烟草野火病发生情况的影响明显,主要表现在病叶率、病情指数、防控效果三个方面。具体结果见图1和表1。3次调查均显示水杨酸处理后的小区,烟草野火病的发病率最小 (见图1)。
由表1分析可知:水杨酸及其他常规植物激素物质在伸根期期施用对野火病的控制效果普遍较差,水杨酸和氨鲜酯具有一定效果分别为36.38%、16.75%;第2次药剂处理后 (7月9日) 调查结果显示,2次施用水杨酸处理后的小区对野火病的控制效果(60.63%) 明显好于对照药剂农用链霉素单独施用一次的效果(31.10%),且优于于常规植物激素物质赤霉素(22.35%)和胺鲜酯(32.48%),但差异不显著(P>0.05);第3次处理后 (7月25日) 调查结果显示,3次使用水杨酸处理对野火病的控制效果更佳(71.90%),明显优于对照药剂农用链霉素施用2次的效果(30.04%),同时也优于常规植物激素物质赤霉素(42.09%)和胺鲜酯(48.74%),且水杨酸处理与其他处理的差异显著(P<0.05)。
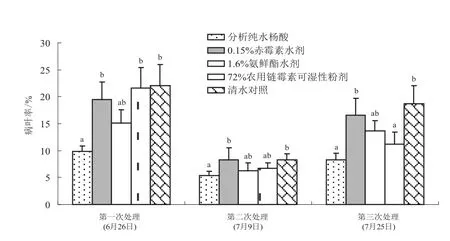
图1 水杨酸对烟草野火病病叶率的影响
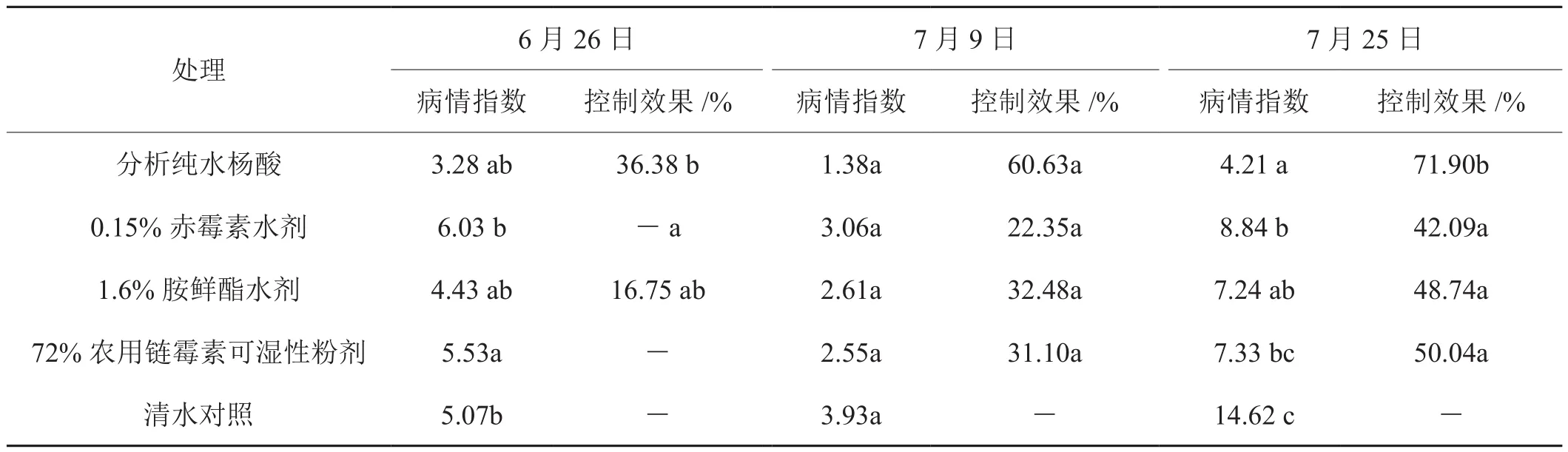
表1 水杨酸对烟草野火病的控制效果
2.2 水杨酸对烟株生物学性状的影响
水杨酸对烟株生物学效应主要体现在有效叶片数、株高、茎围以及最大叶面积四个指标上。具体结果见图2至图5。
有效叶片数(图2)团棵期 (6月27日) 各处差异不明显,在10.20~11.13之间,打顶期(7月26日) 处理2 (赤霉素) 对烟株的有效叶片数有明显的抑制作用,而水杨酸及其他处理对叶片数的影响较小。处理2 (赤霉素) 对株高(图3)有促进作用,且这种作用在打顶期较为明显,水杨酸及其他处理对烟草株高的影响较小。处理2 (赤霉素) 对烟株茎围(图4)有明显的抑制作用,水杨酸及其他处理与空白对照差异很小。赤霉素的早期施用对烟株叶面积(图5)的增加有明显的抑制作用,胺鲜酯处理能促进了烟株叶面积的增加,水杨酸与其他处理对烟草叶面积的影响较小。
总之,在叶片数、株高、茎围以及最大叶面积上,水杨酸与空白对照相比不存在较大的促进作用,也不存在副作用。
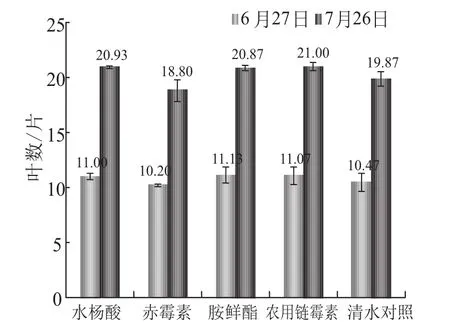
图2 不同处理对烟株叶片数的影响
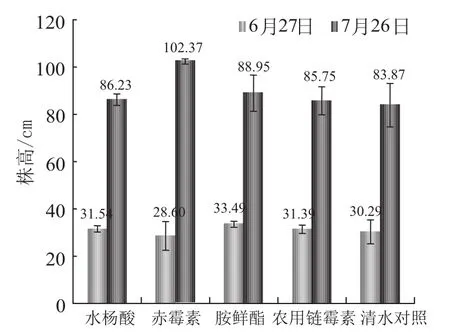
图3 不同处理对对烟株株高变化的影响

图4 不同处理对烟株茎围变化的影响
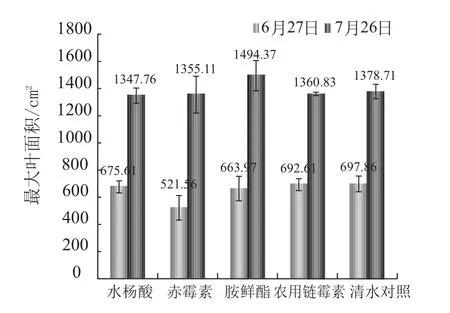
图5 不同处理对烟株最大叶面积的影响
3 讨论
从水杨酸诱导烟草抵抗野火病的效果上看,水杨酸通过提前处理增加烟草自身抗病能力对烟草抵抗野火病的效果,明显好于在发病后用农用链霉素连续处理2次直接防治野火病的效果,这一结论与其他研究者就水杨酸在烟草或其他作物上关于抵抗病害的研究结论相一致:如郝中娜等[13]测定了7种非杀菌剂对烟草野火病病菌的室内抑菌效果及其作用烟株以后的诱抗效果,结果表明:7种物质对野火病菌均没有直接的杀伤作用,但水杨酸和草酸作用于烟株以后对野火病具有较好的诱抗效果;王媛等[14]对拟南芥植株喷施水杨酸后,接种灰霉病菌(Botrytis cinerea),观察植物对病原物反应的变化,结果表明:在平板培养中水杨酸对灰霉菌分生孢子的萌发及菌丝的生长均无明显抑制作用,但对拟南芥植株用水杨酸处理后导致植株体内PAL和POD活性升高、木质素积累从而产生抗病性,病情明显减少。近年来,大量的研究[15-20]证明,水杨酸在激活双子叶植物的防御反应中起着重要的作用。
水杨酸作用于烟草能诱导其抵抗野火病,且具有较好效果,这种效果的产生可能通过以下几种方式实现:(1)改变植物抗病性酶活性。水杨酸一方面能够增加超氧化物歧化酶(SOD)等促进H2O2产生酶类的活性,另一方面能够抑制过氧化氢酶(CAT)、抗坏血酸过氧化物酶(APX)等促进H2O2降解酶的活性[21],从而促进植物体内超氧阴离子(·O2-)、羟自由基(·OH)和过氧化氢(H2O2)的含量的增加,这些物质可以直接杀伤病原物,参与膜脂过氧化,介入超敏反应(HR),同时促进细胞壁木质化和HR细胞的死亡,使感病植物获得抗病性。(2)诱导病程相关蛋白(PR蛋白)的合成。PR蛋白是在植物受到病原物侵染以后产生的一种或多种蛋白质,它具有抗蛋白水解酶的特性,可以抵抗病原菌的侵染,是植物潜在的抗性物质。用水杨酸处理植物可以诱导某些PR蛋白的产生,从而使植株产生系统抗病性[22-24]。如用水杨酸处理烟草可以诱导烟株产生9种PR蛋白的mRNA,并在转录过程中调节PR蛋白合成[25];Ohashi用SA处理烟草叶片7天后,PR蛋白的含量高达100 μg/gFW,是未用水杨酸处理叶片的1000多倍,在如此大量PR蛋白的组织中TMV病毒都难以存活[26]。(3)抑制病原致病性酶活性。病原侵染后能够分泌多聚半乳糖醛酸酶、蛋白酶、纤维素酶等用来降解寄主细胞壁的胞外酶,实现对寄主植物的侵染与破坏,但用水杨酸处理植物以后可以诱导植物产生抑制这些降解酶活性的物质,实现对植物的保护。如Palva研究[27]发现,水杨酸对细菌Erwinia carotovorasubsp. Carotovora分泌的植物细胞壁降解酶具有抑制作用,用1 mmol/L的水杨酸处理烟草叶片后,细菌的生长就被抑制,处理4~5天后,抑制效果最显著。
水杨酸作为一种植物调控物质,其在生产使用上具有使用剂量小、使用时间要求严格、使用环境要求高等特点,因此,在使用过程中要特别注意其准确的使用浓度、适当的处理部位、正确的处理时间等,才能保证其诱导作物抵抗病害的效果。另外,水杨酸水溶性较小、只溶于有机溶剂,将其作为防治烟草野火病的有效物质,在以后的应用和研究过程中要进一步加快以水杨酸为有效成分的新型诱导杀菌剂的研制与开发工作,不断完善其用于防治烟草野火病的最佳施用时间和剂量的筛选,并针对烟草整个生育期组配关于野火病控制的系统控制方案。
4 结论
水杨酸作为一种新型的植物激素物质,对烟草野火病的控制具有一定效果,且无毒副作用,符合未来绿色、安全、优质烟叶生产的需要,具有巨大的潜在发展空间。
[1]魏代福, 谭青涛, 张广民. 防治烟草野火病的药剂测定与筛选[J]. 中国烟草科学, 2010, 31(2): 42-44.
[2]王振国, 丁伟. 烟草野火病发生与防治的研究进展[J].中国烟草学报, 2012, 18(2): 101-106.
[3]刘雅婷, 张世珖, 李永忠, 等. 防治烟草野火病的药剂筛选及应用研究[J]. 湖南农业大学学报, 2002, 28(2): 109-111.
[4]原永兵, 曹宗巽. 水杨酸在植物体内的作用[J]. 植物学通报, 1994, 11(3): 1-9.
[5]李德红, 潘瑞炽. 水杨酸在植物体内的作用[J]. 植物生理学通讯, 1995, 31(2): 144-149.
[6]Raskin I. Salicylate, a new plant hormone [J]. Plant Physiology, 1992, 99(3): 799-803.
[7]Daw B D, Zhang L H, Wang Z Z. Salicylic acid enhances antifungal resistance to Magnaporthe grisea in rice plants[J].Australasian Plant Pathology, 2008, 37: 637-644.
[8]张智慧, 聂燕芳, 何磊, 等. 外源水杨酸诱导水稻相关防御酶活性及内源水杨酸含量的变化[J]. 华中农业大学学报, 2010, 29(5): 541-545.
[9]张衍荣, 王小菁, 张晓云, 等. 水杨酸对豇豆枯萎茵的抑制作用[J]. 华中农业大学学报, 2006, 25(6): 610-613.
[10]王霞, 王振中, 纪春燕, 等. 水杨酸诱导水稻抗菌物质对稻瘟病的抑制作用[J]. 华中农业大学学报, 2011, 30(2):193-196.
[11]谢鑫, 桑维军, 张新强, 等. 水杨酸诱导烟草抗TMV作用的研究[J]. 江苏农业科学, 2011, 39(2): 151-153.
[12]杜小凤, 吴传万, 杨文飞. 等. 寡聚糖和水杨酸对黄瓜根结线虫的防治[J]. 中国农学通报, 2011, 27(10): 280-283.
[13]郝中娜, 文景芝, 李永镐, 等. 非杀菌性化学物质诱导烟草抗野火病的初步研究[J]. 烟草科技, 2000, (7):45-46.
[14]王媛, 杨红玉, 程在全. SA诱导拟南芥对灰霉病的抗性与木质素含量的关系[J]. 植物保护, 2007, 33(4):50-54.
[15]Malamy J, Henning J, Klessig D F. Temperature depended induction of salicylic acid and its conjugates during the resistance response to tobacco mosaic virus infection [J].Plant Cell, 1992, 4(3): 359-366.
[16]Rasmussen J B, Hammerschmidt R, Zook M N. Systemic induction of salicylic acid accumulation in cucumber after inoculation with Pseudomonas syringae pv. Syringae [J].Plant physiol, 1991, 97(4): 1342-1347.
[17]Shah J, Tsui F, Klessig D F. Characterization of a salicylic acidinsensitive mutant (sail) of Arabidopsis thaliana, identi fied in a selective screen utilizing the SA-inducible expression of the lms2 gene [J]. Mol Plant Microbe Interact, 1997, 10: 69-78.
[18]Bowling S A, Guo A, Cao H, et al. A mutation in Arabidopsis that leads to constitutive expression of systemic acquired resistance [J]. Plant Cell, 1994, 6: 1845-1857.
[19]Antoniw J F, White R F. The effects of asoirin and polyacrylic acid on soluble leaf proteins and tesistance to virus infection in five cultivars of tobacco[J]. Phytopathol Z,1980, 98(4): 331-341.
[20]Thibaud-Nissen F, Wu H, Richmond T, et al. Development of Arabidopsis wholegenome micro arrays and their application to the discovery of binding sites for the TGA2 transcription factor in salicylic acid-treated plants [J]. Plant Journal, 2006, 47(1): 152-162.
[21]Bowler C, Alliote T. The induction of mangangese superoxide dismutase in response to stress in Nicotiana plumbaginifolia [J]. The EMBO Journal, 1989, 8(1): 31-38.
[22]原永兵, 刘成连, 鞠志国. 水杨酸对苹果叶片中过氧化氢的调节及其机制[J]. 园艺学报, 1997, 24(3): 220-224.
[23]Okuno T, Nakyama M, Okajima N, et al. Systemic resistance to downy mildew and appearance of acid soluble proteins in cucumber leaves treated with biotic and abiotic inducers [J]. Ann Phytopath Soc Japan, 1991, 57: 202-211.
[24]Reglinski T, Poole P R, Whitakr G, et al. Induced resistance against Sclerotina sclerotorum in kiwifruit leaves [J]. Plant Pathology, 1997, 46: 716-721.
[25]Bol J F, Van Kan J A. The synthesis and possible functions of virus-induced proteins in plants [J]. Microbiol Sci, 1988,5(2):47-52.
[26]Ohashi Y, Matsuoka M. Localization of pathogenesis related proteins in the epidemis and intercellular space of tobacco mosaic virus infection [J]. Plant Cell Physiol, 1987, 28(7): 1227-1235.
[27]Palva T K, Hurtig M, Saindrenan P, et al. Salicylic acid induced resistance to Erwinia carotovora subsp. carotovora in tobacco [J]. Molecular Plant-Microbe Interactions, 1994,7(3): 356-363.
猜你喜欢
科学大众(2024年5期)2024-03-06 09:39:50
江西农业(2022年2期)2022-02-25 04:09:26
成都信息工程大学学报(2018年4期)2019-01-23 06:57:20
中国烟草学报(2016年1期)2016-11-16 08:38:11
西南农业学报(2016年4期)2016-05-17 05:42:12
中国药物应用与监测(2015年5期)2015-12-11 03:15:53
化工进展(2015年6期)2015-11-13 00:27:14
结核与肺部疾病杂志(2015年3期)2015-07-18 11:08:56
应用化工(2014年11期)2014-08-16 15:59:13
中国烟草学报(2012年2期)2012-04-09 06:45:18
