
SCoT分子标记技术初步应用于烟草属部分材料的遗传分析及种间杂种的鉴定
2013-03-14 08:35:48刘超党江波魏烨昕吴天姣汪卫星郭启高梁国鲁
中国烟草学报 2013年5期
刘超,党江波,魏烨昕,吴天姣,汪卫星,郭启高,梁国鲁
西南大学园艺园林学院,重庆 400716
目标起始密码子多态性(Start codon targeted polymorphism,SCoT)标记是Collard和Mackill[1]于2009年发明的一种基于SPAR目的基因分子标记, 是一种能跟踪性状的分子标记技术。与传统分子标记相比,具有易于建立体系、与性状连锁、重复性较好、物种间通用等特点,近年来得到一定程度的应用。该分子标记最早应用于水稻[1],其后,在菠萝[2]、葡萄[3]、龙眼[4]、草莓[5]、柑橘[6]、牡丹[7]中也得到应用。目前,已应用于烟草遗传分析的分子标记有RAPD,SSR,ISSR,AFLP,SRAP等[8],但SCoT应用于烟草中还未见相关报道。
烟草生产病害严重,亟待优良抗病品种[9]。在烟草种及同属野生种中存在大量抗病资源,将具抗病性的野生种与栽培品种进行杂交一直是烟草获得抗病力的主要方法之一[10]。在杂交育种中,对杂种后代的早期鉴定是非常重要的环节。目前,尚未见利用SCoT标记进行种间杂种鉴定的报道。因此,本文利用SCoT标记技术对烟草种间杂种进行鉴定,为烟草分子标记辅助育种提供参考。
1 材料与方法
1.1 试验材料
供试材料共23份,包括两个野生种N. glutinosa、N. plumbaginifolia和11个烟草品种Beinhart1000-1(雪茄),K326(烤烟),L-8(白肋),Coker176(烤烟),Florida301(雪茄),TI448A(白肋),遵烟6号(烤烟),长征1号(烤烟),云烟85(烤烟),云烟97(烤烟),云烟87(烤烟),见表1。N. plumbaginifolia作父本与云烟87杂交后代10株,分别编号为YP1、YP2…YP10。均种植于西南大学果树学重点实验室基地。

表1 供试材料
36个SCoT引物序列SP1、SP2……SP36均参照Collard和Mackill[1],由上海生工生物工程有限公司合成。DL2000 marker,dNTPs,Taq DNA聚合酶,10×Buffer等均购自宝生物工程(大连)有限公司。
1.2 试验方法
1.2.1 DNA提取及PCR反应
DNA提取采用改良CTAB法[11]。
PCR反应体系及反应程序参照韩国辉等[12]的报道反应体系为20μL: 10×buffer 2.0μL,Mg2+1.875mmol/L,dNTP 0.25mmol/L,Taq DNA聚合酶2.5Μ,的DNA模版50 ng,引物0.75μmol/L。反应程序为: 94℃预变性4min,94℃变性1min,50℃退火1min,72℃延伸2min,循环36次,72℃延伸10min。
电泳及拍照:扩增反应结束后,采用8%的聚丙烯酰胺凝胶电泳,使用银染检测。
1.2.2 条带记录与数据统计分析
SCoT分子标记技术是多位点显性标记技术,对扩增清晰且易于辨认的条带采用0-1系统记录其位置,在相同迁移位置有带的记为“1”,无带记为“0”,建立SCoT标记的0、1矩阵。利用NTSYS-2.0软件,把得到的0-1矩阵输入到NTSYS-2.0计算遗传相似系数并按ΜPGMA方法进行聚类分析。
2 结果与分析
2.1 SCoT分子标记在烟草中的适用性
以“云烟97”为模板,分别用36个SCoT引物进行扩增,扩增产物的电泳图谱见图1,各引物所扩增出的条带数介于3-15,大部分引物扩增出的条带较多,也较清晰(图1)。 可见该体系适于SCoT引物对烟草基因组DNA的扩增。

图1 36条引物扩增云烟97总DNA所得产物的电泳图谱
2.2 不同烟草材料的SCoT分子标记多态性
2.2.1 11个烟草品种的SCoT分子标记多态性
筛选9条多态性较好的引物对11个烟草品种进行扩增,结果发现引物Sp15在Beinhart1000-1、L-8、Florida 301、TI448A、CZ-1中均扩增出长度约750 bp的条带,这5个烟草品种除CZ-1为烤烟外,其余均非烤烟;SP30在K326、Coker176、云烟85、云烟87、云烟97这5个烤烟品种中扩增出一条长度约500 bp的条带,而其余品种未见此条带。此外,亦有Sp21在L-8、Florida 301、TI448A、云烟85中扩增出一条约750bp的条带,Sp27在Beinhart1000-1、L-8、Florida 301、TI448A、CZ-1扩增出了一条约750bp的条带。Sp6在Florida 301、TI448A中扩增出约750bp的条带,在其他品种中没有扩增出该条带。这与袁洪[13]得出的白肋烟和烤烟品种遗传多样性指数存在显著差异相一致。祁建民等[14]认为烟草栽培种经自然和人工选择,产生了种内不同类型和较大的遗传变异。
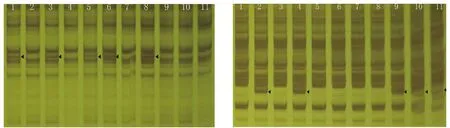
图2 引物Sp-15(A)和引物Sp-30(B)对普通烟草的各种类型DNA扩增出的条带
分析SCoT引物在普通烟草的各种类型普通烟草的各种类型扩增的条带多样性较差。9条引物, 共扩增出条带118条,其中多态性条带仅42条,多态性比例仅35.59%。经相似性分析表明,各烟草品种间的相似性系数均高于0.9,而传统标记如ISSR等技术分析普通烟草的各种类型相似系数介于0.6-1.0[15-16]。可见SCoT引物扩增出的多样性条带比例较低,不宜用于普通烟草的遗传关系分析,但引物Sp-15、Sp-27、Sp-30可在一定程度上区分某一类型(如烤烟)与其它类型品种。这可能与SCoT不能覆盖全基因组有关。
2.2.2 3个烟草种间SCoT分子标记的多态性
应用上面的SCoT体系对N. glutinosa、N.plumbaginifolia与普通烟草Beinhart1000-1进行扩增发现。9条引物共扩增出163条带,其中多态性条带135条,多态性比例为82.82%,其中SP27号引物的多态性比例最高,达到了90.91%,说明3个烟草种的多态性比较丰富。相似性分析表明,N. glutinosa与11个烟草品种的相似系数在0.5左右,与梁景霞等[16]的报道相近;N. plumbagini flia与11个烟草品种的相似系数在0.36-0.39,与李凤霞等[17]的报道相近。可见,SCoT标记适于上述3个烟草种遗传关系的分析。

图3 引物SP3(A)、SP13(B)、SP23(C)、SP24(D)、SP34(E)扩增N. glutinosa(1)、N. plumbaginifolia(2)、Beinhart1000-1(3)3个材料DNA产物的电泳图谱
2.3 烟草远缘杂交后代群体的SCoT鉴定
从36条SCoT引物中筛选出能在两亲本间扩增出特异条带的引物17条,其中17条均能在子代中扩增出母本特异条带,17条中有11条能在子代中扩增出父本的特异条带。据统计,子代具有的与母本相同的特异条带有76条,与父本相同的特异条带48条,可见10株杂交后代都为云烟87与蓝茉莉叶烟草的真杂种,且这10株杂种与其母本云烟87更为相似。
图4是引物SP6的扩增结果,从图中可看出,10个子代均能扩增出2条父本特异条带和3条母本特异条带。

图4 引物SP-6对亲本和10株杂种后代扩增结果注: 所示为父本特异条带;所示为母本特异条带。
上述能在子代中扩增出父本的特异条带的11条引物能在10株或部分后代中扩增出父母本特异条带的引物所扩增出的特异条带数量有一定差异,部分引物如SP8、SP13、SP27、SP30、SP34能在部分后代中扩增出父母本所没有的特异条带。
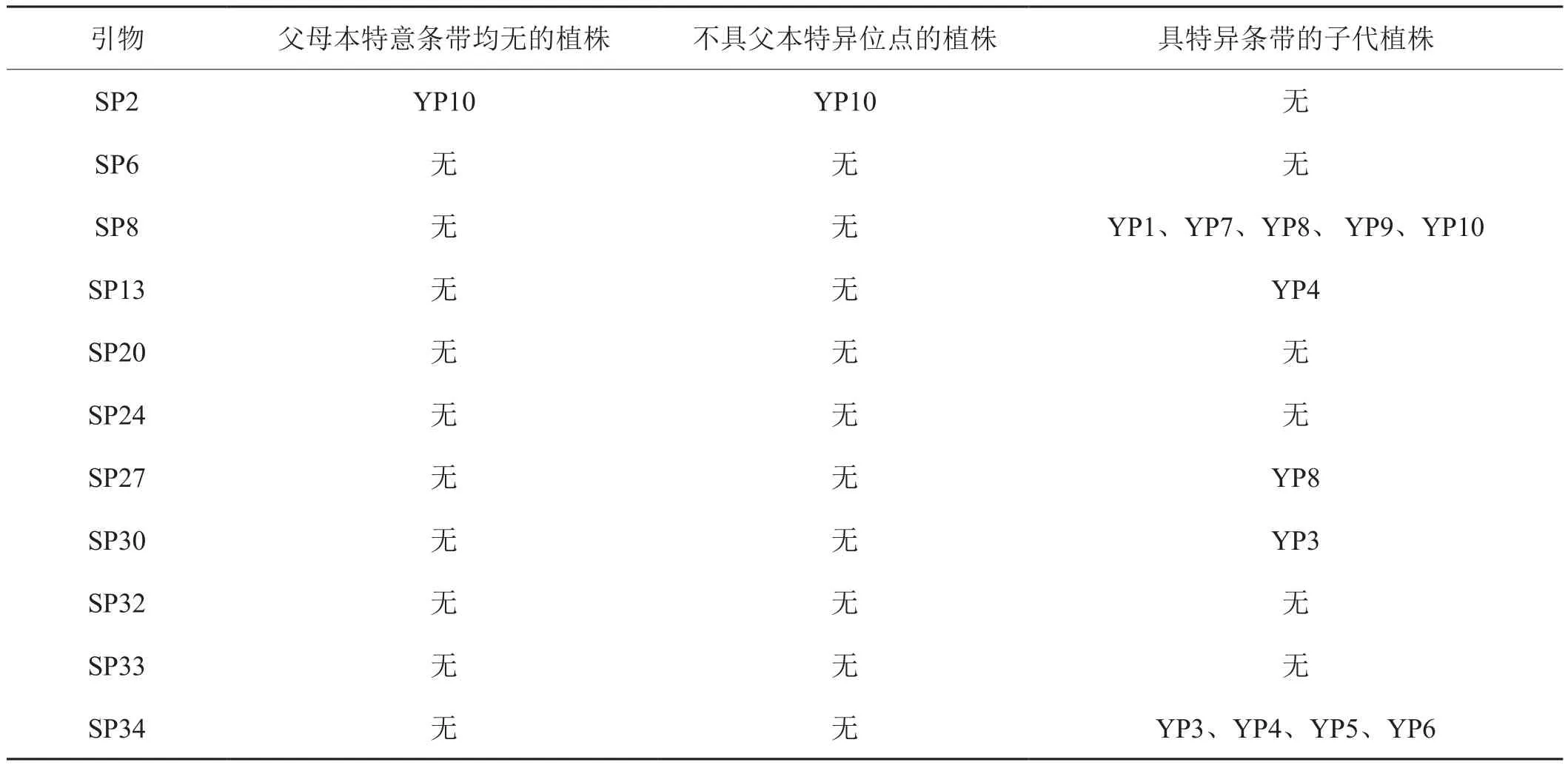
表2 11条引物对10株后代扩增情况
3 讨论
目标起始密码子多态性利用的是启动密码子(ATG)两端碱基的保守性和多态性设计而成,其扩增产物因启动子两端碱基的变化以及碱基插入-缺失引起的变化而呈现不同的片段,这说明该引物可跟踪性状变化[18]。SCoT标记与功能基因相关,能对部分与性状相关的DNA序列进行差异扩增,因此在同一种不同品种间的多态性并不是非常丰富,但其所示差别可能与相关性状相关[19]。但该标记并不能覆盖全基因组,其多态性可能不能与其它标记相比。在本研究中,普通烟草的各种类型间的多态性较差,但是部分引物可以将部分烤烟品种与晒烟、白肋烟相区分开来,这为烟草性状的基因定位提供了一定的参考。由于烟草部分种间性状差异较大,如本研究所用的黏烟草和蓝茉莉叶烟草,其在植株形态、花的形态等外观形态方面均存在巨大差异,因此通过SCoT标记可以将其区分开来。
远缘杂交是现代作物育种中的一个重要方面。烟草属中众多野生种是烟草栽培品种的优良抗源,将野生种的抗性转入普通烟草中是目前烟草抗病育种的一个重点和难点,其中远缘杂交是一个常用的方法[10]。在远缘杂交中,对后代是否真杂种的鉴定非常重要,常规的杂种鉴定主要有形态学和细胞学的方法。杂种后代苗期形态差异不大,所以形态学鉴定较为费时。细胞学的鉴定则需通过染色体核型分析等手段,由于染色体制片的繁琐及其对实验技术的高要求,对于部分杂种后代的鉴定也较为费时[20]。因此,选用分子标记进行杂种鉴定较为理想[21]。贾志刚等[22]运用SCoT分子标记分析了枇杷杂交后代与亲本之间的多态性。韩国辉等[6]也将SCoT标记应用于沙田柚杂交后代的群体鉴定。本研究中成功的应用SCoT分子标记鉴定了10株云烟97与蓝茉莉叶烟草的杂交后代的杂种真实性。
由于作物育种主要对其性状进行关注,因此,与性状密切连锁的分子标记对加快育种进程有重大意义。将SCoT分子标记应用于烟草材料的遗传分析将对烟草的分子标记辅助育种作重要补充。
[1]Bertrand C Y Collard, David J Mackill. Start codon targeted(SCoT) polymorphism: a simple, novel DNA marker technique for generating gene-targeted markers in Plants [J].Plant Mol Biol Rep ,2009,27(1):86-93.
[2]陈香玲,苏伟强,刘业强,等.36份菠萝种质的遗传多样性Scot分析[J].西南农业学报,2012,25(2):625-629.
[3]张君玉,郭大龙,龚莹,等.葡萄目标起始密码子多态性反应体系的优化[J].果树学报,2011,28(2):209-214.
[4]陈虎,何新华,罗聪,等.龙眼24个品种的SCoT 遗传多样性分析[J].园艺学报,2010,37(10):1651-1654.
[5]秦国新,何桥,梁国鲁,等.草莓属植物Scot分析体系的建立及优化[J].果树学报,2012,29(3):393-397.
[6]韩国辉,向素琼,汪卫星,等.柑橘SCoT分子标记技术体系的建立及其在遗传分析中的应用[J].园艺学报,2011,38(7):1243-1250.
[7]侯小改,王娟,贾甜,等.牡丹SCoT分子标记正交优化及引物筛选[J].华北农学报,2011,26(5):92-96.
[8]徐双红,吴成林,戴林建,等.烟草遗传标记研究进展[J].Crop Research, 2011, 25(3): 272-276.
[9]王刚.我国烟草病虫害防治研究策略探讨[J].中国烟草科学,2003, (4):37-39.
[10]杨铁钊. 烟草育种学[M]. 北京:中国农业出版社,2003.
[11]梁景霞, 祁建民, 吴为人,等. 烟草DNA的提取与SRAP反应体系的建立[J]. 中国烟草学报, 2005, 11(4):33-38.
[12]韩国辉,汪卫星,向素琼,等.多倍体枇杷Scot分析体系的建立与优化[J].果树学报,2011,28(3):433-437.
[13]袁洪.烟草种质资源遗传多样性分析及地方种质资源评价[D].中国,重庆,西南大学,2012.
[14]祁建民,王涛,陈顺辉,等.部分烟草种质遗传多样性与亲缘关系的ISSR标记分析[J].作物学报,2006,32(3):373-378.
[15]肖炳光,杨本超. 利用 ISSR 标记分析烟草种质的遗传多样性[J]. 中国农业科学 2007, 40(10): 2153-2161.
[16]梁景霞,祁建民,方平平,等.烟草种质资源遗传多样性与亲缘关系的I SSR聚类分析[J].中国农业科学,2008, 41(1):286-294.
[17]李凤霞, 王卫锋, 王鲁,等.烟草属植物遗传多样性和亲缘进化关系的荧光AFLP 分析[J]. 中国农业科学,2010, 43(12): 2418-2427.
[18]熊发前, 唐荣华, 陈忠良,等.目标起始密码子多态性(SCoT):一种基于翻译起始位点的目的基因标记新技术[J]. 分子植物育种, 2009, 7(3): 635-638.
[19]熊发前, 唐荣华, 陈忠良,等. 目标起始密码子多态性(SCoT)分子标记技术在花生属中的应用[J]. 作物学报.2010, 36(12): 2055-206.
[20]Anna TROJAK-GOLUCH, Apoloniusz BERBEE.Cytological investigations of the interspecific hybrids of Nicotiana tabacum L. × N. glauca Grah [J]. J Appl Genet,2003, 44(1): 45-54.
[21]乔燕春,林顺权,何小龙,等.普通枇杷种内和种间杂种苗的RAPD鉴定[J].果树学报,2010,27(3):385-390.
[22]贾志刚.三倍体批把实生后代及杂交后代SCOT标记及DNA甲基化分析[D]. 重庆:西南大学,2011.
猜你喜欢
农业技术与装备(2020年10期)2020-11-03 11:25:44
系统工程与电子技术(2016年2期)2016-04-16 05:16:53
浙江柑橘(2016年3期)2016-03-11 20:12:48
河北书画研究(2016年6期)2016-02-03 02:57:44
中国光学(2015年1期)2015-06-06 18:30:20
诗歌月刊(2014年3期)2015-04-14 17:50:51
传奇故事(破茧成蝶)(2015年8期)2015-02-28 09:29:41
温州医科大学学报(2014年7期)2014-07-18 02:43:24
食品工业科技(2014年15期)2014-03-11 18:17:30
海岸工程(2014年4期)2014-02-27 12:51:28
