
纳米抗体的特性及其应用研究进展
2013-03-03 10:32张宇昊
食品科学 2013年3期
郭 婷,张宇昊,2,马 良,3,*
(1.西南大学食品科学学院,重庆 400716;2.重庆市特色食品工程技术研究中心,重庆 400716;3.农业部农产品贮藏保鲜质量安全风险评估实验室(重庆),重庆 400716)
纳米抗体的特性及其应用研究进展
郭 婷1,张宇昊1,2,马 良1,3,*
(1.西南大学食品科学学院,重庆 400716;2.重庆市特色食品工程技术研究中心,重庆 400716;3.农业部农产品贮藏保鲜质量安全风险评估实验室(重庆),重庆 400716)
纳米抗体与传统IgG抗体相比,具有分子质量小、更容易被生产、稳定性好、抗原结合力高等特点,目前在医学、食品科学等很多方面展开了积极有效的研究和应用。本文重点就纳米抗体的结构、特性和在食品科学领域的应用进行综述和分析,并对纳米抗体在食品检测方面的应用前景进行展望。
纳米抗体;特性;食品;应用
基于抗原-抗体特异性反应的免疫分析技术因其操作简单、准确性高、检测速度快等特点,近些年发展迅猛,在食品安全检测领域中发挥着越来越重要的作用。目前免疫分析法主要利用单克隆抗体、多克隆抗体等来实现真菌毒素、农兽药残留、食品添加剂、非法添加物等各种食品危害物的检测。单克隆抗体的特异性强,容易纯化、标记。但其制备复杂、价格较昂贵[1-2],不能进行沉淀和凝胶反应[1-3],应用范围受到一定限制。多克隆抗体的反应强度大,与抗体的亲和力和灵敏度相对较高,但特异性相对较低,在检测中很容易出现假阳性现象。目前,小分子功能抗体作为免疫学研究的热点[3],主要有Fab片段(L和Fd)、Fv(VH和VL)、ScFv、纳米抗体等。与其他小分子功能抗体相比,纳米抗体的亲和力高、稳定性好,更易制备成稳定的融合子,有较大的发展潜力。
生物学家Ward在1989年分离检测骆驼血清中抗体时发现,骆驼血液中除常规的四链抗体外,还有一些仅含重链的抗体。1993年,Hamers-Casterman等[4]研究表明,骆驼抗体天然缺失轻链,只含有重链,因此又称重链抗体(heavy chain antibodies,HCAbs)。另有研究者[5]发现,骆驼抗体中还缺少CH1。克隆重链抗体的可变区得到只由一个重链可变区组成的单域抗体,称为VHH抗体(variable domain of heavy chain of heavy-chain antibody,VHH)。VHH晶体直径2.5nm,长4nm,因此又称为纳米抗体(nanobody,Nb)[6-7],是自然存在的可与抗原结合的最小片段。纳米粒子具有的颗粒尺寸小、比表面积大、表面能高等特点,以及表面效应、尺寸效应和宏观量子隧道效应[8]使得纳米抗体在诸多方面均优于传统抗体。纳米抗体在疾病的检测和治疗方面有显著应用成果[9-11],而在食品领域的应用处于起步阶段,今后将在食品工业及检测方面发挥巨大的作用。本文主要针对纳米抗体的特性和在食品科学领域中的应用进行综述和分析,对纳米抗体在食品检测方面的应用前景进行展望。
1 纳米抗体的结构
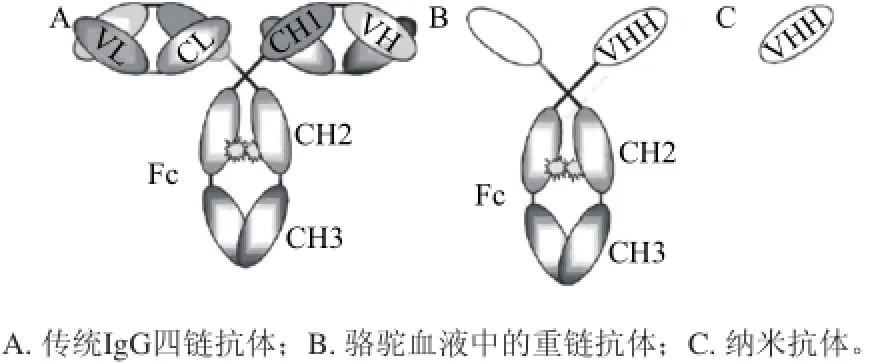
图1 抗体结构模式图[7]Fig.1 Antibody structure[7]
纳米抗体(图1C)一般结构呈椭圆形,体积很小,分子质量为单克隆抗体的1/10(15kD)[12],与普通抗体(图1A)相比,化学性质更加活泼,具有纳物质的量级亲和力,能更有效地与抗原缝隙结合[13]。纳米抗体的基因组中存在恒定区(constant region of heavy chain 1,CH1),它是轻链锚定的部位,但在mRNA形成中被剪切掉,所以纳米抗体缺乏轻链[5]。纳米抗体的二级结构是2个β片层形成支架,3个高变区聚集在一侧参与抗原识别。与骆驼的重链抗体相比,纳米抗体仅由重链可变区组成。而骆驼的重链抗体(图1B)含有一个铰链区,两个恒定区(CH2和CH3)和独特的重链可变区(VHH)。
骆驼源VHH(即Nb)和人体VH的基因同源性达到80%~90%[14],结构十分相似,都包含3个高变区(hypervariable region,HVR)和4个骨架区(framework region,FR)。通过比较发现,骆驼VHH 与人体VH之间存在微小但不容忽视的区别,主要包含两方面:1)它们的互补决定区(complementarity determining region,CDR)不同。VHH的CDR1和CDR3比VH更长。VHH的CDR3长度为16~18个氨基酸,而人和小鼠VH的平均长度分别只有14和12个氨基酸[15-17]。2)它们的区别在于FR2中的氨基酸[18]。普通抗体的FR2中V37、G44、L45和W47这4个氨基酸残基是疏水性残基,在进化中是相当保守的[15]。而VHH中,它们突变为亲水性的氨基酸残基F37、E44、R45、G47,增加了VHH的溶解性[15](图2)。因此,将人源抗体VH结构域FR2中的一些氨基酸进行VHH特征性改造,可以获得稳定性好、溶解性好,并且保持原有抗体特异性和亲和力的VH 抗体[19]。目前,人源化已经取得成功[14]。

图2 人源VH和骆驼源VHH基因序列[7]Fig.2 Gene sequences of human VH and camel VHH[7]
2 纳米抗体的特性
2.1 理化性
纳米抗体在许多理化性质方面具有三方面的明显优势:1)Nb溶解性高。由于Nb的FR2中一些亲水性氨基酸取代疏水性氨基酸,使Nb的溶解性增加,从而提高作为药物的利用率。2)Nb抗热性强。研究发现,在37℃放置1周后Nb仍能保持80%以上的生物活性,这使其在室温条件下更易于使用和保存[20-21]。然而单链抗体的稳定性差,常常显示聚集倾向,在37℃产生聚集现象[22],大大限制了其应用。Nb内部存在二硫键是其抗热性强的主要原因[23]。3) Nb易复性。在高于90℃的环境中长期放置,Nb仍能重新获得生物活性[24-25]。同时,Nb在强变性剂的条件下也表现出较高的耐受性[26]。其他抗体则发生不可逆的热聚合[18]。2.2 抗原结合性
纳米抗体有较高的抗原结合力和目标亲和力。Nb包含了骨架区的氨基酸残基,在一定程度上弥补缺失轻链造成的抗原结合力低的不足;同时Nb的CDR3较长,形成稳定的凸形结构,可深入抗原内部更好地结合抗原。普通抗体只能识别抗原表面的位点。因此Nb的抗原结合能力更广泛,不仅可结合小分子半抗原和肽,还可结合大分子蛋白和病毒,甚至当识别位点被深入包埋时,小分子的Nb也可对其进行表位识别[27]。
2.3 免疫原性
纳米抗体对人体的免疫原性弱,生物相容性好。免疫原性是指能够刺激机体形成特异性抗体或致敏淋巴细胞的能力。免疫原性与分子大小、化学结构等有关,分子质量越小免疫原性越小。Nb分子质量很小,只有一个结构域,缺少Fc段,从而避免了Fc段引起的补体反应,对人体的免疫原性很低。在动物实验中,纳米抗体未引起任何体液和细胞免疫应答[28]。因此,可通过带有人VH FR2烙印的纳米抗体来获得非免疫原性纳米抗体。但有研究表明,纳米抗体作为药物长期反复使用会增加免疫原性,影响治疗[29]。
2.4 穿透性
相对单克隆抗体显影剂组织穿透力差、不易清除的不足[30],纳米抗体的组织穿透能力很强,可以进入致密的组织,并且多余未结合的纳米抗体能够被快速清除,有利于疾病的诊断。Cortez-Retamozo等[31]制备的纳米抗体显影剂穿透性好、亲和力高、肿瘤组织的显像效果好,并且对正常组织几乎没有损伤。近期报道[32],纳米抗体用于分子显像会强烈地滞留在肾脏中,影响胰脏附近检测分子信号的灵敏度。用18F标记的纳米抗体的肾脏滞留量最小(<7%)[33]。另有研究[34]表明,纳米抗体能够有效地穿透血脑屏障,为脑部供药提供新方法。
2.5 生产性
纳米抗体的分子质量小、结构简单,能被单个基因编码,利用基因工程能在酵母菌、大肠杆菌等微生物中大量表达。纳米抗体相对价格低廉、可进行大规模生产,易于普及和应用。涂追等[35]利用半巢式PCR法构建天然噬菌体单域重链抗体文库,文库的多样性较好,可以用于后续淘汰。Ablynx公司表示,他们将酵母反应器酿造纳米抗体的产量提高,达到0.5g/L的产量[31]。
3 纳米抗体在食品科学领域的应用
纳米抗体具有普通抗体没有的独特性质,对其进行基因改造,使其携带特定的结构,在疾病的诊断与治疗上比其他抗体更有优势,近几年在这些方面的应用报道逐年增多。而纳米抗体在食品科学领域,如食品毒理学研究、食品分析检测等方面的研究和应用较晚,但已成为新的研究热点。尤其是近两年涌现出不少研究和应用成果,显示出纳米抗体在该领域应用的巨大潜力。
3.1 食物中毒的解毒
食品毒理学研究表明,Nb能与细菌结合,抑制细菌产酶的活性,从而起到一定的治疗作用。此外,Nb还可以中和细菌毒素、蛇蝎毒素。Hmila等[36]筛选出抗蝎子毒素Aa2hI’的Nb,这种抗体可以高效中和AahI’毒素,用于解毒。
3.2 食品污染物的脱毒
研究表明,应用纳米粒子的表面效应可对饲料中AFT进行脱毒处理,能有效降低AFT的危害[37]。纳米抗体是一种纳米粒子,具有纳米粒子的三大效应,即表面效应、尺寸效应、宏观隧道量子效应。因此可以运用纳米抗体的表面效应进行食品及原料中污染物的脱毒研究。
3.3 食品检测
纳米抗体属于新型抗体,在国内外食品检测方面研究和应用相对较晚,尤其是国内相关的研究和应用报道较少。但随着纳米抗体等小分子功能抗体自身优势不断体现,近些年研究逐渐增多,2010—2011年国内相关研究机构相继报道了一批有关小分子功能抗体的研究结果,充分显示了纳米抗体等小分子功能抗体巨大的应用前景。
据报道[38-39]江南大学制备出抗黄曲霉毒素B1的小分子功能抗体ScFv片段,利用该ScFv片段检测酱油中的黄曲霉毒素B1
[40]。管笛等[41]制备出AFM1标准品的无毒替代物F(ab')片段,也属于小分子功能抗体,并将其用于花生样品检测。另有学者制备出各种小分子功能抗体,如抗克伦特罗ScFv片段[42-43]、抗有机磷ScFv片段[44]、抗氨基甲酸酯ScFv片段[45]等,这些片段可用于建立针对小分子物质的非竞争性免疫检测,具有分子质量小、穿透力强、免疫原性低等优点。然而,有研究[22,46]发现上述片段仍存在稳定性相对较差、亲和性较低等缺点,限制了其在食品检测方面的应用。
2009年,Kirchhofer等[47]筛选出能调节绿色荧光蛋白(GFP)构象及光谱性质的纳米抗体。名为Enhancer的纳米抗体与GFP结合,使荧光效果增强5倍;而名为Minimizer的纳米抗体使蛋白的某特定区远离发光区,从而使荧光效果降低5倍。该研究表明,Nb可以在体内或体外控制蛋白构象。目前,研究人员正在筛选与其他荧光蛋白特异的纳米抗体。利用纳米抗体增强荧光强度来提高食品检测的灵敏性,有重要的现实意义。另外,Muyldermans等[8]重组Nb,将其作为生物传感器的探针。在食品工业中可利用探针能与待测物质特异性结合的性质来检测食品中的有毒有害物质。
4 结 语
纳米抗体的稳定性好、亲和力较高,克服了小分子功能抗体的缺点,同时又具有分子质量小、免疫原性弱、组织穿透力强等单克隆抗体、多克隆抗体不具备的优点,适应用于食品检测、疾病治疗等方面。在食品免疫及相关检测方面还有待于进一步深入地研究和应用,经过综述与分析,在以下3个领域极具开发和研究潜力:1)利用细胞工程技术、基因工程技术等新型技术,构建基于纳米抗体的双功能抗体,与两种性质不同的抗原发生结合,在多残留检测、天然噬菌体单域重链抗体文库构建等方面发挥重要的作用。2)利用纳米抗体对组织穿透力强和对靶位点的亲和力高的特点,进入致密组织,检测被深入包埋的物质,实现肉制品、蛋制品、乳制品、饼粕饲料等各种复杂基质食品中非法添加物和小分子污染物等的高灵敏度检测。3)利用纳米抗体控制荧光蛋白的荧光特性,选择性增强或猝灭荧光,用荧光探针技术提高毒素、农兽药残留物等物质的分析检测灵敏度。
[1] 吴石金, 孙培龙. 简明免疫学原理[M]. 北京: 化学工业出版社, 2008: 173.
[2] 冯仁青, 郭振泉. 现代抗体技术及其应用[M]. 北京: 北京大学出版社, 2006: 55-76.
[3] 胡圣尧. 免疫学与生物技术[M]. 北京: 人民卫生出版社, 2003: 61.
[4] HAMERS-CASTEMAN C, ATARHOUCH T, MUYLDEMANS S, et al. Naturally occurring antibodies devoid of light chains [J]. Nature, 1993, 363: 446-448.
[5] NGUYEN V K, HAMERS R, WYNS L, et al. Loss of splice consensus signal is responsible for the removal of the entire CH1 domain of the functional camel IGG2A heavy-chain antibodies[J]. Mol Immunol, 1999, 36(8): 515-524.
[6] 杨珂, 王冬. 纳米抗体及其应用[J]. 细胞与分子免疫学杂志, 2008(4): 425-427.
[7] MUYLDERMANS S, BARAL T N, CORTEZ RETAMOZZO V, et al.Camelid immunoglobulins and nanobody technology[J]. Veterinary Immunology and Immunopathology, 2009, 128: 178-183.
[8] 李国秀, 李建科. 纳米技术在食品领域中的应用[J]. 粮食与油脂, 2007(8): 13.
[9] SKOTTRUP P D, LEONARD P, KAZMAREK J Z, et al. Diagnostic evaluation of a nanobody with picomolar affinity toward the protease RgpB from Porphyromonas gingivalis[J]. Analytical Biochemistry, 2011, 415: 158-167.
[10] ABBADY A Q, al-MARIRI A, ZARKAWI M, et al. Evaluation of a nanobody phage display library constructed from a Bruella-immunised camel[J]. Veterinary Immunology and Immunopathology, 2011, 142: 49-56.
[11] STEYAERT J, KOBILA K. Nanobody stabilization of G proteincoupled receptor conformational states[J]. Current Opinion in Structural Biology, 2011, 21: 567-572.
[12] MUYLDERMANS S, LAUWEREYS M. Unique single-domain antigen binding fragments derived from naturally occurring camel heavy-chain antibodies[J]. Mol Recognit, 1999, 12: 1-10.
[13] de GENST E, SILENCE K, DECANNIERE K, et al. Molecular basis for the preferential cleft recognition by dromedary heavy-chain antibodies[J]. PNAS, 2006, 103: 4586-4591.
[14] VINCKE C, LORIS R, SAERENS D, et al. General strategy to humanize a camelid single-domain antibody and identification of a universal humanized nanobody scaffold[J]. J Bio Chem, 2009, 284: 3273-3284.
[15] MUYLDERMANS S, ATARHOUCH T, SALDANHA J, et al. Sequence and structure of VH domain from naturally occurring camel heavy chain immunoglobulins lacking light chains[J]. Protein Eng, 1994, 7(9): 1129-1135.
[16] VU K B, GHAHROUDI M A, WYNS L et al. Comparison of llama VH sequences from conventional and heavy chain antibodies[J]. Mol Immunol, 1997, 34: 1121-1131.
[17] WU T T, JOHNSON G, KABAT E A. Length distribution of CDR H3 in antibodies[J]. Proteins: Struct Funct Genetics, 1993, 16: 1-7.
[18] NGUYEN V K, DESMYTER A, MUYLDERMANS S, et al. Functional heavy-chain antibodies in Camelidae[J]. Adv Immunol, 2001, 79: 261-296.
[19] DAVIES J, RIECHMANN L. Camelising human antibody fragments: NMR studies on VH domains[J]. FEBS Letters, 1994, 339(3): 285-290.
[20] GHAHROUDI M A, DESMYTER A, WYNS L, et a1. Selection and identification of single domain antibody fragments from camel heavychain antibodies[J]. FEBS Lett, 1997, 414: 521-526.
[21] van der LINDEN R H, FRENKEN LG, de GEUS B, et al. Comparison of physical chemical properties of llama VHH antibody fragments and mouse monoclonaI antibodies[J]. Biochim Biophys Acta, 1999, 1431: 37-46.
[22] 秦海燕, 毛晓燕, 乔玉玲, 等. 单链抗体的研究进展[J]. 现代生物医学进展, 2011, 11(4): 795-797.
[23] 王更如, 姜广水. 纳米抗体的研究进展[J]. 现代免疫学, 2010, 30(1): 73-77.
[24] PÉREZ J M J, RENISIO J G, PROMPERS J J, et al. Thermal unfolding of a llama antibody fragment: a two-state reversible process[J]. Biochemistry, 2001, 40: 74-83.
[25] EWERT S, CAMBILLAU C, CONRATH K, et al. Biophysical properties of camelid VHH domains compared to those of human VH3 domains[J]. Biochemistry, 2002, 41: 3628-3636.
[26] DUMOULIN M, COURATH K, van MEIRHAEQHE A, et al. Single domain antibody fragments with high conformational stability[J]. Protein Sci, 2002, 11(3): 500-515.
[27] STIJLEMANS B, CONRATH K, CORTEZ-RETAMOZO V, et al. Efficient targeting of conserved cryptic epitopes of infectious agents by single-domain antibodies. African trypanosomes as a paradigm[J]. J Biol Chem, 2004, 279(2): 1256-1261.
[28] CORTEZ-RETAMOZO V, BACKMANN N, SENTER P D, et al. Efficient cancer therapy with a nanobody-based conjugate[J]. Cancer Res, 2004, 64(8): 2853-2857.
[29] 苏幼红, 李江伟. 骆驼来源单域抗体在免疫治疗中的研究进展[J].生物技术通报, 2010(6): 27-32.
[30] 王兴华, 于晓玲, 董宝玮. 超声介导超声造影剂在分子影像与基因治疗领域中的应用[J]. 中华超声影像学杂志, 2004, 13(11): 865-867.
[31] CORTEZ-RETAMOZO V, LAUWEREYS M, HASSANZADEH G H G, et a1. Efficient tumor targeting by single-domain antibody fragments of camels[J]. Int J Cancer, 2002, 98: 456-462.
[32] ANEYCKEN I, D’HUYVETTER M, HERNOT S, et al. Immunoimaging using nanobodies[J]. Current Opinion in Biotechnology, 2011, 22: 877-881.
[33] VANEYCKEN I, XAVIER C, BLYKERS A, et al. Synthesis and first in vivo evaluation of18F-anti-HER2-nanobodies: a new probe for PET imaging of HER2 expression in breast cancer[J]. J Nucl Med, 2011, 52: 664-1664.
[34] HARMSEN M M, de HAARD H J. Properties, production, and applications of camelid single-domain antibody fragments[J]. Appl Microbiol Biotechnol, 2007, 77: 13-22.
[35] 涂追, 许杨, 何庆华, 等. 半巢式PCR法构建天然噬菌体单域重链抗体文库[J]. 食品科学, 2010, 31(19): 299-303.
[36] HMILA I, ABDALLAH RBA, SAERENS D, et al. VHH, bivalent domains and chimeric heavy chain-only antibodies with high neutralizing efficacy for scorpion toxin AahI’[J]. Mol Immunol, 2008, 45: 3847-3856.
[37] 汤俊琪, 庞广昌. 免疫传感器测定食品中的黄曲霉毒素的研究进展[J]. 食品科学, 2009, 30(17): 318-329.
[38] 王铁斌, 丁虎生, 杨炼, 等. 抗黄曲霉毒素B1单链抗体的筛选和鉴定[J]. 微生物学报, 2009, 49(1): 135-140.
[39] 杨炼, 刘自琴, 刘蓉, 等. 抗黄曲霉毒素B1单链抗体的表达载体的比较[J]. 食品科学, 2010, 31(9): 171-176.
[40] 刘蓉, 杨炼, 孙秀兰, 等. 应用抗黄曲霉毒素单链抗体检测酱油中黄曲霉毒素B1[J]. 食品工业科技, 2011, 32(1): 281-283.
[41] 管笛, 李培武, 张奇, 等. 黄曲霉毒素B1标准替代物的制备及其在花生样品检测中的应用[J]. 中国油料作物学报, 2011, 33(5): 503-506.
[42] 王弘, 刘细霞, 潘科, 等. 抗克伦特罗单链抗体基因构建及蛋白结构模拟[J]. 食品科学, 2009, 30(13): 227-231.
[43] 刘细霞, 孙远明, 董洁娴, 等. 抗克伦特罗核糖体展示单链抗体文库的构建及鉴定[J]. 食品科学, 2011, 32(15): 200-204.
[44] 贺江, 梁颖, 樊明涛, 等. 噬菌体展示技术制备甲氧基有机磷农药抗独特型抗体[J]. 分析化学研究报告, 2011, 39(2): 178-182.
[45] 王俊平, 张伟伟, 杜欣军, 等. 西维因单链抗体基因克隆、表达及活性分析[J]. 食品工业科技, 2010, 31(11): 161-164.
[46] 齐永华, 董永军, 宁红梅, 等. 单链抗体技术在农兽药残留检测方面的研究进展[J]. 东北农业大学学报, 2011, 49(9): 7-11.
[47] KIRCHHOFER A, HELMA J, SCHMIDTHALS K, et al. Modulation of protein properties in living cells using nanobodies[J]. Nat Struct Mol Biol, 2009, 17: 133-138.
Properties and Application Prospects of Nano-Antibody in Food Industry
GUO Ting1,ZHANG Yu-hao1,2,MA Liang1,3,*
(1. College of Food Science, Southwest University, Chongqing 400716, China;2. Food Engineering and Technology Research Center of Chongqing, Chongqing 400716, China;3. Laboratory of Quality and Safety Risk Assessment for Agro-products on Storage and Preservations (Chongqing), Ministry of Agriculture, Chongqing 400716, China)
Compared with IgG antibodies, nano-antibodies have the properties with smaller size, easier production, high stability and excellent aff i nity. Currently, nano-antibodies are developed and applied in the fi eld of medical and food science. In this paper, the structure, properties and applications of nano-antibodies in food industry have been reviewed and analyzed, which will provide extensive application prospect in food detection.
nano-antibody;property;food;application
TS207.3
A
1002-6630(2013)03-0294-04
2011-12-28
国家“863”计划项目(2007AA10Z427);中央高校基本科研业务费专项(XDJK2011B001)
郭婷(1988—),女,硕士研究生,研究方向为食品安全与质量控制。E-mail:heavencandy@163.com
*通信作者:马良(1979—),女,副教授,博士,研究方向为食品安全与食品检测技术。E-mail:zhyhml@163.com
猜你喜欢
考试与评价·高二版(2021年3期)2021-09-10
数学物理学报(2020年5期)2020-11-26
天然产物研究与开发(2018年8期)2018-09-10
中国比较医学杂志(2018年6期)2018-06-26
科教导刊(2017年10期)2017-06-09
上海农业学报(2017年4期)2017-04-10
心血管病防治知识(2016年9期)2016-11-21
公民与法治(2016年14期)2016-05-17
特产研究(2016年3期)2016-04-12
华南农业大学学报(2015年5期)2015-12-04
