
外源NO对Pb胁迫下亚麻幼苗的保护效应
2013-03-03 01:32:18李宇洁
甘肃农业科技 2013年11期
李宇洁,安 梅
(兰州职业技术学院,甘肃 兰州 730070)
植物生长过程中,常常会遭受到重金属离子如铅、汞、镉、砷、锌等的污染[1~3]。铅是一种具有对植物强毒性的重金属,铅土壤污染是当今世界面临的主要环境问题之一[4],土壤中的铅被植物吸收后,可导致植物中毒,特别是导致植物根部、植株等形态生理变化[5]。亚麻(Linseed)在逆境胁迫下的生理生化指标变化已有大量报道,如水、盐、干旱、镉等协迫[6~9],但关于Pb污染方面的研究未发现报道。一氧化氮(NO)在植物光态建成、生长发育、衰老等过程和对非生物胁迫的响应中有重要作用[10]。有研究表明,NO可以缓解非生物胁迫下植物氧化损伤,其效应与NO处理浓度有关[11~12],低浓度的NO能够缓解盐胁迫对燕麦和番茄叶片的氧化性损伤,能够有效提高镉离子胁迫下亚麻幼苗叶片自我保护效应[6,13~14]。我们研究Pb胁迫与不同浓度SNP(NO供体)处理下亚麻的生理生化指标变化,旨在探讨Pb胁迫下亚麻防御机理以及NO在植物逆境中的作用。
1 材料与方法
1.1 供试材料
指示亚麻品种为陇亚10号。
1.2 试验方法
试验共5个处理,处理A,对照(CK);处理B,0.2mmol/LPb(Ac)2;处理C,0.1mmol/LSNP+0.2 mmol/L Pb(Ac)2;处理D,0.5 mmol/L SNP+0.2 mmol/L Pb(Ac)2;处理E,1.0 mmol/L SNP+0.2 mmol/LPb(Ac)2。3次重复。
挑选饱满的亚麻种子,用蒸馏水冲洗浸泡12 h,再用0.1%升汞溶液消毒30 s,然后播种育苗。幼苗长至4片叶后移栽到装有已灭菌蛭石的花盆,每盆30株。每隔2 d浇1/8浓度的Hogland营养液100 mL,保持湿度在60%~70%。生长35 d后,选取生长整齐一致、健康的亚麻植株按试验设计进行SNP预处理,即分别量取50mL不同浓度SNP溶液喷施于叶面,每隔24 h处理1次,连续处理3次。第3次处理后24 h,在蛭石中施加有0.2m mol/LPb(Ac)2的Hogland营养液100mL,每隔24 h处理1次,连续处理4次。施Pb(Ac)2前及处理后24、48、72、96 h采集生长状况一致的幼苗叶片,取0.5 g幼苗叶片,用茚三酮显色法测定游离脯氨酸(Pro)含量[15],用硫代巴比妥酸法测丙二醛(MDA)含量[16]。再取各处理幼苗叶片0.5 g,加入pH 7.8的0.05mol/L磷酸缓冲液(含1%PVP)研磨成匀浆,定容至10mL,在4℃、10 000 rpm下离心20min,取清液备用[17]。分别用NBT显色法测定过氧化氢酶(CAT)活性[18]、用愈创木酚氧化法测定过氧化物酶(POD)活性[19]、紫外吸收法测定超氧化物歧化酶(SOD)的活性[20]。
数据采用Excel 2003整理,使用SPSS 18.0分析。
2 结果与分析
2.1 Pb胁迫下SNP处理对亚麻幼苗中脯氨酸(Pro)含量的影响
逆境条件能够诱发植物脯氨酸含量积累。胁迫开始时各处理亚麻幼苗叶片内Pro含量无显著差异(P>0.05)。随着胁迫时间的延长,处理B、D、E逐渐升高(图1),均在96 h后Pro含量达到最大值;而处理C却随着胁迫时间的延长呈先升后降再升高的趋势,在96 h时达到最大值。在胁迫的各个时间段,SNP处理的Pro含量均低于处理B,说明SNP处理可以缓解Pb胁迫下植物氧化损伤,其中处理C效果最明显,Pro含量较B处理在48、72与96 h时分别降低了44.42%、30.12%、29.86%(P<0.01)。可知,SNP处理可以显著缓解叶片氧化损伤,降低Pro的积累速度。
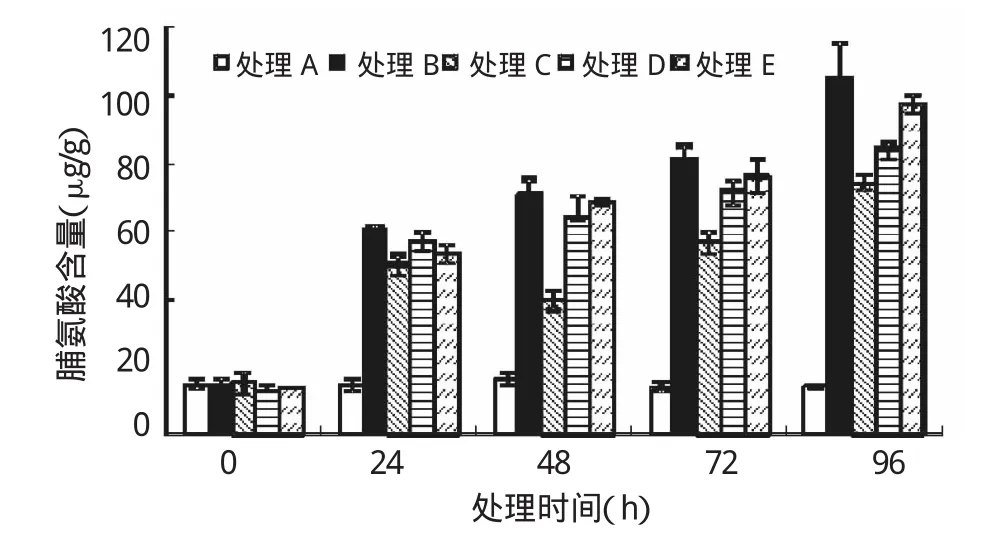
图1 外源NO对Pb胁迫下亚麻幼苗中脯氨酸的影响
2.2 Pb胁迫下SNP处理对亚麻幼苗中丙二醛(MDA)含量的影响
由图2可知,胁迫开始时各处理含量无显著差异(P>0.05)。随胁迫时间的延长,处理B、E叶片内MDA含量随之逐渐升高,在96 h时达到最大值,与对照差异极显著(P<0.01),较对照提高190.64%、159.15%;处理C、D中MDA含量逐渐升高,至72 h时达到最大值,与对照差异极显著(P<0.01),较对照提高136.88%、140.92%,而后随之降低。SNP处理在胁迫各时间段,其MDA含量均低于单纯Pb胁迫(处理B)。至96时处理C、D、E的MDA含量较处理B降低了44.21%(P<0.01)、13.35%(P<0.05)、12.15%(P<0.05),其中以处理C效果最佳。由此看出,SNP能够缓解Pb协迫下亚麻幼苗叶片中MDA含量的升高。
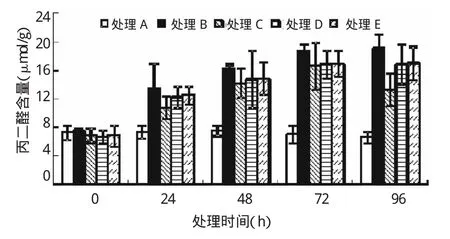
图2 外源NO对Pb胁迫下亚麻幼苗中丙二醛的影响
2.3 Pb胁迫下SNP处理对亚麻幼苗中抗氧化酶(SOD、CAT和POD)活性的影响
由图3可知,在Pb胁迫96 h内,SNP对亚麻幼苗SOD、CAT和POD活性具有显著的影响。Pb胁迫开始时,SNP各处理CAT活性均高于单纯胁迫(处理B)。随着Pb胁迫时间的延长,处理B、C、D、E中CAT活性呈先升高后逐渐降低的趋势,在24 h时CAT活性达到最大值,均高于对照处理,其中以处理C效果显著(P<0.05),较处理B提高了14.30%;24 h后,各处理CAT活性均快速下降,处理B下降程度较处理C、D、E快,至96 h达到最低值,均低于对照处理,其中以处理C的CAT活性下降速率最慢。可知,外源SNP能够降低Pb胁迫幼苗叶片CAT活性的下降速率,提高CAT活性,以处理C效果最为显著。
胁迫初期,SNP处理POD活性均高于对照。随着胁迫时间的延长,POD活性呈先升高后下降的趋势(图3b)。在48 h时,Pb、SNP处理组POD活性达到最大值,其中处理C的POD活性最大,较对照提高了122.04%(P<0.01);随着胁迫时间延长,各处理组POD活性均下降,其中处理B的POD活性下降速率最大,在96 h降至最低,显著低于对照(P<0.05)和处理C(P<0.01)。说明外源NO可提高亚麻幼苗逆境下POD活性。
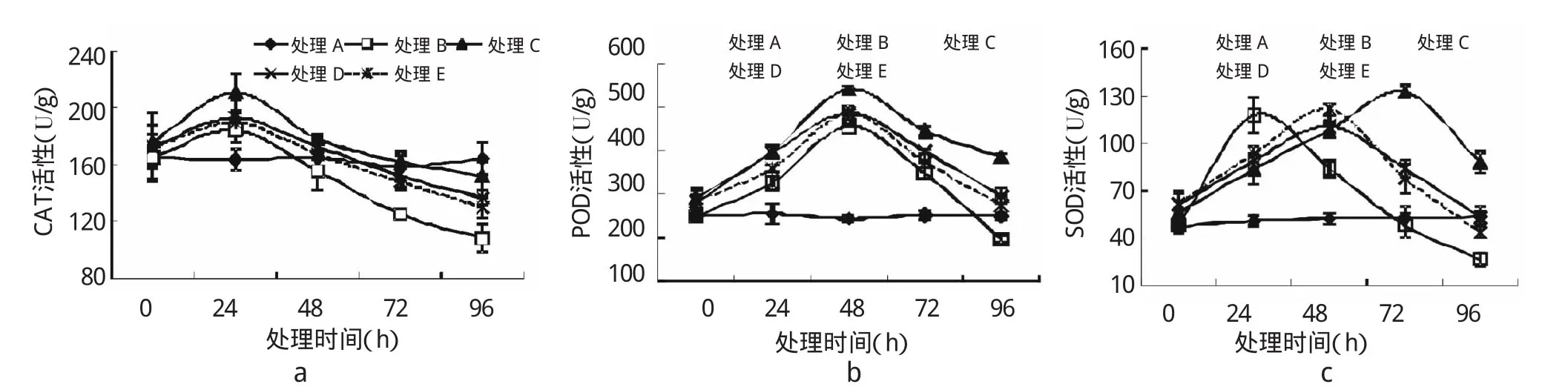
图3 外源NO对Pb胁迫下亚麻幼苗中SOD、CAT、POD酶活性的影响
由图3c可知,在胁迫初期,SNP各处理SOD活性均高于对照;随着胁迫时间的延长,处理B、C、D、E的SOD活性呈先升高后下降的趋势。其中处理B在24 h时SOD活性达到最大值;处理D、E在48 h达到最大值;处理C在72 h时达到最大值,说明SNP处理可缓解亚麻幼苗超氧阴离子的生成。其后降低过程中处理B中SOD活性下降速率最快,处理C最慢,说明处理C效果最佳。
3 小结与讨论
1) 研究结果表明,NO缓解了Pb胁迫下亚麻幼苗叶片中丙二醛含量的升高,促进脯氨酸含量升高,提高了过氧化氢酶、过氧化物酶、超氧化物歧化酶等抗氧化酶的活性,减缓抗氧化酶的活性Pb胁迫下的下降速率,缓解叶片受到的氧化损伤。且保护效应与SNP的浓度相关,0.1mmol/LSNP处理效果显著优于1.0mmol/LSNP处理。即NO具有清除植物体内活性氧,促进抗逆性物质的积累,调节抗氧化酶活性的功能。
2)植物在逆境胁迫时,植物体内会产生大量的活性氧,若逆境时间过长就会导致细胞受到氧化损伤,引发细胞膜脂质过氧化反应,导致细胞膜裂解和丙二醛含量增加。本研究对丙二醛含量测定的结果说明,外源NO对细胞膜具有良好的保护作用,可减轻Pb胁迫对其造成的伤害,这与王宪叶等人外源NO能够缓解渗透胁迫下的小麦幼苗叶片膜脂过氧化,提高其抗性的结论吻合[21]。
3)植物体内脯氨酸的积累在一定程度上反映了植物的抗逆性。本研究表明,外源NO能促进Pb胁迫下亚麻幼苗叶片中游离脯氨酸含量的积累,这与NO不仅可以提高黑麦草的抗冷性和番茄的抗盐性[14,22],还能增强了亚麻的抗重金属Pb能力,以及李源等人在蚕豆上取得的结果相吻合[23]。
4)Pb胁迫对这3种抗氧化酶的诱导效果并不一致,这可能与Pb胁迫过程中SOD、POD和CAT在膜脂质过氧化过程所起的不同作用有关,如SOD可以清除超氧阴离子,其产物是过氧化氢,过氧化氢含量的增加,导致了CAT失活。同时,H2O2和O2-有可能与CAT形成复合物,抑制了CAT的活性[24]。
[1]夏增禄, 穆从如,孟维奇,等.Cd、Zn、Pb及其相互作用对烟草小麦的影响[J].生态学报,1984,4(3):231-236.
[2]安建平,焦成瑾,王廷璞,等.Cd2+胁迫对黄瓜幼苗生物膜损伤和保护酶的影响[J].甘肃农业科技,2006(4):4-7.
[3]任继凯,陈清朗,陈灵芝,等.土壤中镉、铅、锌及其相互作用对作物的影响[J].植物生态学与地植物学丛刊,1982,6(4):320-329.
[4]SHEN ZG,LIX D,WANG CC,et al.Lead phytoextraction from contaminated soil with high-biomass plant species[J].Environ.Qual.,2002,31:1893-1900.
[5]杨元根,PATERSON E,CAMPBELL C.城市土壤中重金属元素的积累及其微生物效应[J].环境科学,2001,22(3):44-48.
[6]徐 严,魏小红,李兵兵.外源NO对重金属Cd胁迫下亚麻幼苗叶片抗氧化能力的影响[J].甘肃农业大学学报,2012,47(5):45-49.
[7]张晓平,薛召东,郝冬梅,等.亚麻耐渍的生理机制研究初探[J].中国麻业科学,2007,29(3):169-172.
[8]王红梅,金忠民,孙雪瑰,等.盐胁迫对亚麻幼苗生长的影响[J]. 齐齐哈尔大学学报,2015,21(4):87-89.
[9]王晓琦,沙 伟,徐忠文.亚麻幼苗对干旱胁迫的生理响应[J]. 作物杂志,2005(2):13-15.
[10]张绪成,上官周平,高世铭.NO对植物生长发育的调控机制[J]. 西北植物学报,2005,25(1):812-818.
[11]BELIGNIM V,LAMATTINA L.Is nitric oxide toxic or protective[J].Trends in Plant Science,1999,4(8):229-300.
[12]BELIGNIM V,LAMATTINA L.Nitric oxide protects against cellular damage produced bymethylviologen herbicides in potato plants[J].Nitric Oxide,1999,3(3):199-202.
[13]苏 桐,龙瑞军,魏小红,等.外源NO对NaCl胁迫下燕麦幼苗氧化损伤的保护作用[J].草业学报,2008,17(5):48-53.
[14]苏 桐,魏小红,丁学智,等.外源NO与蔗糖对盐胁迫下番茄幼苗氧化损伤的保护效应[J].生态学报, 2008,28(4):1558-1564.
[15]赵福庚,刘友良.大麦幼苗多胺合成比脯氨酸合成对盐胁迫更敏感[J]. 植物生理学报,2000,26(4):243-349.
[16]赵世杰,许长成,邹 琦,等.植物组织中丙二醛测定方法的改进[J]. 植物生理学通讯,1994,30(3):207-210.
[17]李合生.植物生理生化实验原理与技术[M].北京:高等教育出版社,2000:260-261.
[18]CHANDER M S.Enzymic associationswith resistance to rust and pow deymildew in pea[J].Indian Journal of Horticulture,1990,47(3):341-345.
[19]孙 群,胡景江.植物生理学研究技术[M].杨凌:西北农林科技大学出版社,2006:113-115.
[20]DHINDSA R S,PLUMB D P,THORPE T A.Leaf senescence correlated with increased levels ofmembrane permeability and lipid peroxidation and decreased levels dismutase and catalase[J].Journal of Experimental Botany,1982,32:91-101.
[21]王宪叶,沈文彪,徐朗莱.外源NO对渗透胁迫下小麦幼苗叶片膜脂过氧化的缓解作用[J].植物生理与分子生物学报,2004,3(4):195-200.
[22]马向丽,魏小红,龙瑞军,等.外源一氧化氮提高黑麦草抗冷性机制的研究[J].生态学报,2005,25(6):1269-1274.
[23]李 源,李金娟,魏小红.镉胁迫下蚕豆幼苗抗氧化能力对外源NO和H2O2的响应[J].草业学报,2009,18(6):186-191.
[24]ASADA K.The water-water cycle in chlorop lasts:scavenging of active oxygens and dissipation of excess photons[J].Annual Review of Plant Physiology and PlantMolecular Biology,1999,50:601-639.
猜你喜欢
纺织科学研究(2023年9期)2023-10-23 11:18:12
中老年保健(2022年3期)2022-11-21 09:40:36
科学与财富(2019年15期)2019-10-21 13:35:48
发酵科技通讯(2018年2期)2018-07-06 11:38:46
畜牧兽医杂志(2018年2期)2018-06-15 02:38:38
中国麻业科学(2018年6期)2018-04-09 11:22:12
纺织科学研究(2017年4期)2017-05-17 04:00:02
湖南林业科技(2017年1期)2017-02-06 05:29:00
纺织服装流行趋势展望(2016年6期)2016-05-04 03:52:43
水生生物学报(2015年1期)2015-02-28 16:00:43
