
粳稻杂种后代胚乳淀粉分支酶基因表达特性分析
2013-02-20 09:21金正勋王露露刘海英徐振华张忠臣
东北农业大学学报 2013年1期
金正勋,王露露,刘海英,徐振华,曲 莹,沈 鹏,张忠臣
(东北农业大学农学院,哈尔滨 150030)
由线性直链淀粉和高度分支支链淀粉组成的淀粉是水稻籽粒中主要的能量贮藏物质,约占糙米干重90%。因此籽粒灌浆过程主要是直链淀粉和支链淀粉的合成与积累过程。茎、叶等源器官制造的光合同化物以蔗糖形式运输到籽粒后,在一系列酶促作用下形成直链淀粉和支链淀粉[1]。参与直链淀粉和支链淀粉合成的酶很多,如蔗糖合成酶、ADPG焦磷酸化酶、UDPG焦磷酸化酶、淀粉合成酶、淀粉分支酶等。其中淀粉分支酶的主要作用是剪切a-1,4糖苷键转化为a-1,6糖苷键而形成葡聚糖链分支点,对稻米直链淀粉和支链淀粉合成与积累影响很大的关键酶之一[1-2]。由于直链淀粉是影响稻米蒸煮食味品质最重要的内在因素之一,所以围绕淀粉分支酶活性与直链淀粉含量和蒸煮食味品质的关系国内外已进行很多研究[3-7]。高等植物SBE基因主要包括SBE1、SBE3和SBE4几个类型,它与作物籽粒中支链淀粉的合成密切相关,其中SBE1和SBE3起主导作用,分别调控约70%和30%的支链淀粉合成[8-9]。SBE基因与直链淀粉和支链淀粉含量的关系已有较多报道[10-14]。品种间有性杂交仍然是目前选育水稻新品种和杂种优势利用的主要方法。品种间有性杂交产生的后代数量性状会产生超亲遗传变异。虽然过去对数量性状遗传规律做了大量研究,但数量性状产生超亲变异的分子机理方面研究甚少。因此,本研究选用以籽粒直链淀粉含量作为选择指标,从F2代起连续定向选择培育成的籽粒直链淀粉含量有显著差异的F10代,比较分析籽粒灌浆过程中亲本和杂种后代籽粒直链淀粉含量积累和淀粉分支酶活性以及淀粉分支酶基因mRNA表达量等变化特点,旨在为水稻杂种后代直链淀粉含量产生超亲遗传变异的分子生化学机理提供理论依据。
1 材料与方法
1.1 材料
在东农423×藤系180(组合Ⅰ)和系选1号×通769(组合Ⅱ)中,以籽粒直链淀粉含量为选择指标,从F2代起按高低方向连续定向选择至F7,从中选用籽粒直链淀粉含量差异大的后代各2个(H4、L11为组合Ⅰ;H16、L20为组合Ⅱ)及亲本,2011年在东北农业大学校内进行盆栽试验,盆规格为直径25 cm,高30 cm。4月10日播种,大棚旱育苗,5月25日插秧,每个盆插生长一致的4棵苗,正常肥水管理。抽穗时选取同一天抽穗进行挂牌标记,待抽穗后第10、17、24天分别取挂牌标记的5个穗,取穗中上部灌浆一致的20粒,剥去颖壳和胚后置于冻存管中-80℃保存备用。
1.2 淀粉含量和分支酶活性测定方法
取上述冻存的籽粒,在程方民等方法基础上改进[15],按照章显光等提出的冷碱液糊化后煮沸的方法测其胚乳直链淀粉含量[16];采用硫酸-蒽酮法测定胚乳总淀粉含量[17];胚乳总淀粉含量与直链淀粉含量差值作为支链淀粉含量;参照李太贵等方法测定胚乳淀粉分支酶活性,其酶活性以OD值表示[3]。
1.3 淀粉分支酶基因表达水平分析
采用Trizol法提取籽粒总RNA,以RNase-free DNaseⅠ处理,消除可能的基因组DNA污染,取纯化后的RNA进行反转录,产物用于RT-PCR。第一链cDNA合成采用Promega公司的M-MLV逆转录试剂盒说明进行。以反转录得到的cDNA为模板,利用淀粉分支酶基因特异引物进行扩增(见表1),扩增产物进行琼脂糖凝胶电泳,然后对扩增谱带进行表达丰度的比对分析。

表1 淀粉分支酶基因引物Table 1 Name of RT-PCR primer and sequence
2 结果与分析
2.1 亲本及杂种后代胚乳直链淀粉积累特性比较
灌浆不同时期亲本及杂种后代胚乳直链淀粉含量多重比较结果见表2。
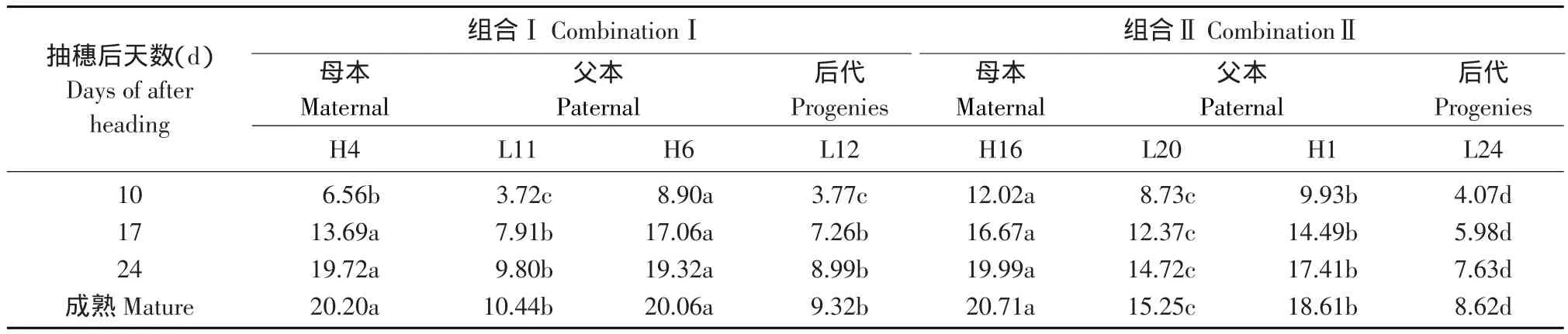
表2 灌浆不同时期胚乳直链淀粉含量变化Table 2 Changes of amylose content in crossing progenies during grain filling (%)
由表2可知,后代直链淀粉含量与双亲比较两个组合都有显著差异,在组合Ⅰ中没有超亲变异,但在组合Ⅱ中有超亲变异,L24的直链淀粉含量显著低于低亲。说明直链淀粉含量选择效果因杂交组合而异,通过连续定向选择可以选育直链淀粉含量超亲的后代。
在灌浆过程中,不同直链淀粉含量亲本及后代胚乳直链淀粉积累趋势基本一致,表现为灌浆前期积累速度快,抽穗17和24 d时积累量分别已达到总直链淀粉含量的67.77%~85.21%和88.50%~97.60%,说明胚乳直链淀粉的合成和积累,主要在灌浆前、中期。
从多重比较结果可知,直链淀粉含量高的亲本及后代在灌浆过程中合成和积累的直链淀粉量显著高于直链淀粉含量低的亲本及后代。说明胚乳直链淀粉合成和积累速度的快慢主要取决于遗传因素,直链淀粉含量高的基因型在灌浆过程中合成和积累的直链淀粉量总是高于直链淀粉含量低的基因型。
2.2 亲本及杂种后代胚乳淀粉分支酶活性比较
灌浆不同时期亲本及杂种后代胚乳淀粉分支酶活性多重比较结果按组合列于表3。
由表3可知,灌浆过程中两个组合直链淀粉含量不同的亲本及后代胚乳淀粉分支酶活性变化趋势基本一致,表现为随灌浆进程酶活性逐渐增加,达到峰值后又逐渐下降,除组合Ⅱ的L20外,都呈单峰曲线变化,峰值都出现在抽穗后第17天。
由灌浆不同时期的两个组合亲本及后代胚乳淀粉分支酶活性多重比较可知,直链淀粉含量高的亲本及后代的酶活性均比直链淀粉含量低的亲本及后代高,其酶活性峰值除组合Ⅰ中的H4与L11无显著差异外,其他都有显著差异,尤其是组合Ⅱ的L24灌浆不同时期的酶活性均显著低于低亲,组合Ⅰ中24 d的H6酶活性显著高于高亲H4。相关分析表明,灌浆不同时期胚乳直链淀粉含量与淀粉分支酶活性间均呈正相关,相关系数分别为0.2568、0.6149、0.5949。说明胚乳直链淀粉含量的合成与积累受淀粉分支酶活性的调控影响较大,通过直链淀粉含量的连续定向选择可以改变杂种后代的淀粉分支酶活性。
2.3 亲本及杂种后代胚乳淀粉分支酶基因mRNA表达量变化动态
灌浆不同时期亲本及杂种后代3种胚乳淀粉分支酶同工型基因OsSBE1、OsSBE3及OsSBE4的mRNA表达量变化动态按组合分别示于图1、2。

表3 灌浆不同时期亲本及后代胚乳淀粉分支酶活性比较Table 3 Comparison of SBE activity in crossing progenies during grain filling(OD·grain-1·min-1)
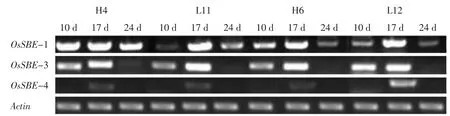
图1 不同灌浆时期亲本及杂种后代胚乳淀粉分支酶基因mRNA表达量变化(组合Ⅰ)Fig.1 mRNA expression of gene involved in SBE on different filling stage(CombinationⅠ)
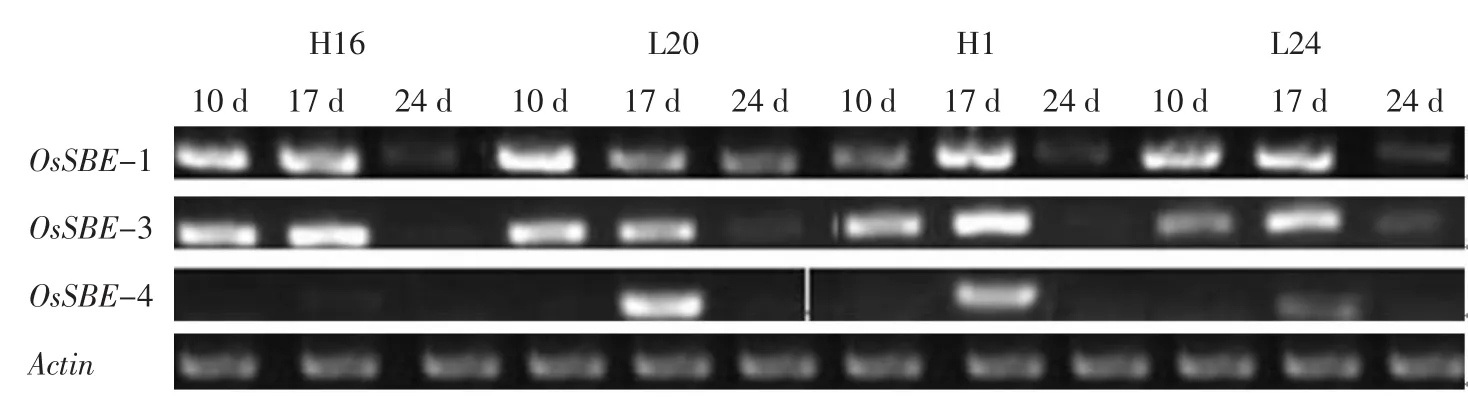
图2 不同灌浆时期亲本及杂种后代胚乳淀粉分支酶基因mRNA表达量变化(组合Ⅱ)Fig.2 mRNA expression of gene involved in SBE on different filling stage(CombinationⅡ)
由图1、2可知,亲本及杂种后代3种胚乳淀粉分支酶同工型基因OsSBE1和OsSBE3的mRNA表达量随灌浆进程逐渐增加,达到峰值后又逐渐下降,呈单峰曲线变化,除组合Ⅰ中L12的OsSBE3外,其余表达量峰值都出现在抽穗后17 d。在灌浆过程中胚乳OsSBE4基因的mRNA表达量相对比较小,除抽穗17 d时能检测到明显的表达量之外,其他时期未检测到明显的表达量。说明胚乳直链淀粉的合成积累主要与OsSBE1和OsSBE3基因关系密切。
2.4 亲本及杂种后代胚乳淀粉分支酶基因mRNA相对表达量比较
在本试验中抽穗后17 d是胚乳淀粉分支酶基因mRNA表达水平最高的时期。为了明确直链淀粉含量高低与胚乳淀粉分支酶基因mRNA表达量间的关系,计算抽穗后17 d的亲本及杂种后代3种胚乳淀粉分支酶同工型基因OsSBE1、OsSBE3及OsSBE4的mRNA相对表达量,结果见表4。

表4 亲本及杂种后代胚乳淀粉分支酶基因mRNA相对表达量比较Table 4 Comparison of SBE gene relative expression level in parent and hybrid progeny
由表4可知,SBE1基因在组合I中,直链淀粉含量高的后代H6与亲本H4、L11相比mRNA表达量无显著差异,但直链淀粉含量低的后代L12比低亲L11显著低,低1.2倍;在组合Ⅱ中,直链淀粉含量高的后代H1与高亲H16相比mRNA表达量无显著差异,但直链淀粉含量低的后代L24与低亲L20相比mRNA表达量显著高,高1.2倍;SBE3基因在组合I和组合Ⅱ中,直链淀粉含量高的后代与高亲、直链淀粉含量低的后代与低亲相比mRNA表达量都没有显著差异;SBE4基因在组合I中,直链淀粉含量高的后代与高亲相比mRNA表达量没有显著差异,但直链淀粉含量低的后代与低亲相比显著低,低1.5倍;在组合Ⅱ中,直链淀粉含量高的后代与高亲、直链淀粉含量低的后代与低亲相比mRNA表达量都显著高。说明根据胚乳直链淀粉含量连续定向选择形成的杂种后代胚乳淀粉分支酶基因的mRNA表达量随着胚乳直链淀粉含量的变化发生变化,而且能超亲表达。由表5可知,胚乳直链淀粉含量与SBE1和SBE3的mRNA表达量呈不显著正相关,与SBE4的mRNA表达量呈显著负相关;胚乳淀粉分支酶活性与SBE1的mRNA表达量呈显著正相关,与SBE3的mRNA表达量呈不显著正相关,与SBE4的mRNA表达量呈不显著负相关。说明直链淀粉含量和淀粉分支酶活性高的基因型其SBE1和SBE3的mRNA表达量也高,低的基因型其mRNA表达量也低,SBE4的表达量与此相反。

表5 胚乳淀粉分支酶基因mRNA相对表达量与直链淀粉含量及淀粉分支酶活性间相关系数Table 5 Correlation analysis of SBE gene relative expression,amylose content and enzymes activities
3 讨论与结论
水稻中SBE3基因缺失表现为直链淀粉含量增加,支链淀粉含量明显降低[10-11]。张鹏利用根癌农杆菌介导的转基因技术将SBE1、SBE3的反义或RNA干扰结构分别导入具有不同直链淀粉含量的水稻受体品种中,结果显示转反义SBE1/SBE3和SBE3-RNAi种子直链淀粉含量较对照都有明显的提高,且不同品种间差异显著[12]。吴方喜等利用农杆菌介导法将水稻SBE1正、反义基因分别导入籼稻恢复系明恢81中,结果表明,转SBE1反义基因的直链淀粉含量明显上升[13]。汪结明等用转基因方法干扰水稻SBE3基因的表达,其结果表明,导入的SBE3基因RNA干扰结构降低目的基因表达,使其SBE活性在籽粒发育各时期均显著降低,且不同株系间具有差异,两个转基因水稻株系各时期籽粒直链淀粉含量均显著高于对照[14]。本试验结果表明,在灌浆过程中籽粒淀粉分支酶活性和SBE基因的mRNA表达量变化呈正相关,且直链淀粉含量和淀粉分支酶活性高基因型其SBE1和SBE3的mRNA表达量高,低基因型其mRNA表达量低,SBE4表达量与此相反。上述国内外研究结果和本试验结果都说明SBE基因对籽粒直链淀粉合成存在调控作用。由于淀粉分支酶的主要作用是剪切a-1,4糖苷键转化为a-1,6糖苷键而形成葡聚糖链分支点[1-2],因此SBE基因对直链淀粉合成积累的调控可能是间接的调控,即通过参与直链淀粉合成相关基因的表达来影响直链淀粉的合成积累。
基因的表达顺序是由DNA转录成mRNA,再以mRNA作为模板合成酶,最终通过各种酶促反应形成表现型性状。籽粒直链淀粉是在AGPase、GBSS、SBE等多种酶参与下合成和积累,是相关基因表达的最终产物。本试验选用的直链淀粉含量高低不同的杂种后代是通过以直链淀粉含量为选择指标,在同一个杂交组合中连续定向选择培育的稳定后代。由本试验结果可知,通过连续定向选择不仅改变杂种后代籽粒直链淀粉含量和淀粉分支酶活性,而且也改变了淀粉分支酶基因的mRNA表达量。在本试验中杂种后代胚乳直链淀粉含量、淀粉分支酶活性、淀粉分支酶基因mRNA表达量等三者的变化是线性同步变化,而且能超亲表达。说明杂种后代胚乳直链淀粉含量的提高与淀粉分支酶活性提高和SBE基因mRNA表达量的增加有密切关系。通过基因终端产物数量的变化可以引起基因mRNA表达量的变化。至于连续定向选择如何引起杂种后代基因mRNA表达量的变化,该变化是否与基因组中基因拷贝数或基因结构的变化有关,有待深入研究。
[1]Reddy K R,Ali S Z,Bhattacharya K R.The fine structureof rice starch amylopectin and its relation to the texture of cooked rice[J].Carbohydr Polym,1993,22:267-275.
[2]高振宇,黄大年,钱前.植物支链淀粉生物合成研究进展[J].植物生理与分子生物学学报,2004,30(5):489-495.
[3]李太贵,沈波,陈能,等.Q酶对水稻籽粒垩白形成的影响[J].作物学报,1997,23(3):338-344.
[4]赵步洪,张文杰,常二华,等.水稻灌浆期籽粒中淀粉合成关键酶的活性变化及其与灌浆速率和蒸煮品质的关系[J].中国农业科学,2004,37(8):1123-1129.
[5]沈鹏,金正勋,罗秋香,等.水稻灌浆过程中籽粒淀粉合成关键酶活性与蒸煮食味品质的关系[J].中国水稻科学,2006,20(1):58-64.
[6]钟连进,程方民.水稻籽粒灌浆过程直链淀粉的积累及其相关酶的品种类型间差异[J].作物学报,2003,29(3):452-456.
[7]Han Y P,Xu M L,Liu X Y,et al.Genes coding for starch branching enzymes are major contributors to starch viscosity characteristics in waxy rice(Oryza sativa L.)[J].Plant Science,2004,166:357-364.
[8]Mizuno K,Kawasaki T,Shimada H.A alteration of the structural properties of starch by the lack of an isoform of starch branching enzyme in rice seeds[J].J Biol Chem,1993,268:19084-19091.
[9]Mizuno K,Kawasaki T,Arai Y.Starch branching enzymes from immature rice seeds[J].J Biochem,1992,112:643-651.
[10]Hiroaki Y,Yasunori N.Organ specificity of isoforms of starch branching enzyme in rice[J].Plant Cell Physiol,1992,33:985-991.
[11]Nishi A,Nakamura Y,Tanaka N.Biochemical and genetic analysis of the effects of amylase extender mutation in rice endosperm[J].Plant Physiol,2001,127:459-472.
[12]张鹏.抑制淀粉分支酶类基因表达对稻米品质影响的研究[D].扬州:扬州大学,2008.
[13]吴方喜,谢华安,苏军,等.RBE1正、反义基因改变籼稻直链淀粉含量的研究[J].福建农业学报,2006,21(2):150-153.
[14]汪结明,张建,江海样,等.RNA干扰水稻SBE3基因的表达对籽粒淀粉合成及其关键酶活性的影响[J].作物学报,2010,36(2):313-320.
[15]程方民,蒋德安,吴平,等.早稻籽粒灌浆过程中淀粉合成酶的变化及温度效应特征[J].作物学报,2001,27(2):201-206.
[16]章显光,黄永楷.稻米直链淀粉含量、糊化温度和胶稠度的初步研究[J].湖北农学院学报,1992,12(1):10-15.
[17]白宝璋,汤学军.植物生理学测试技术[M].北京:中国科学技术出版社,1993.
猜你喜欢
中国糖料(2021年3期)2021-07-13
辽宁农业科学(2021年1期)2021-03-17
生物加工过程(2020年3期)2020-06-17
中国粮油学报(2019年4期)2019-07-12
热带农业科技(2019年1期)2019-01-14
中国果业信息(2019年11期)2019-01-05
小学阅读指南·高年级版(2016年9期)2016-10-31
中国塑料(2016年2期)2016-06-15
中国粮油学报(2016年5期)2016-01-23
新疆农垦科技(2014年10期)2014-02-28
