
择伐强度对天然云冷杉林空间分布格局的影响
2013-01-04 09:34亢新刚
中南林业科技大学学报 2013年1期
沈 林,杨 华,亢新刚,岳 刚
( 北京林业大学 省部共建森林培育与保护教育部重点实验室,北京100083)
择伐强度对天然云冷杉林空间分布格局的影响
沈 林,杨 华,亢新刚,岳 刚
( 北京林业大学 省部共建森林培育与保护教育部重点实验室,北京100083)
探讨不同强度的择伐对天然云冷杉林空间分布格局的影响,为合理经营森林提供依据。采用点格局分析法的O-ring统计,对天然云冷杉林在3种不同强度(弱度20%、中度30%、强度40%)择伐22年后主要树种的空间分布格局及树种间空间关联性进行了分析。结果表明:4块样地林分整体和主要树种均表现为随机分布,强度择伐林木多呈聚集分布,而弱度、中度择伐树种大多处于随机和均匀分布;主要树种间的空间关联性表现为弱度择伐的负相关现象多于其它择伐类型。弱度择伐更加有利于天然云冷杉林林分空间结构的优化。
天然云冷杉林;择伐强度;空间分布格局;O-ring统计
森林空间结构特征是识别森林生态系统健康与稳定的最重要信息[1]。林木的空间分布格局反映了林地内微环境、气候和光照因子、竞争及林木生物学特性等综合作用的结果,对林分内各种空间关系的描述和解释成为了国际上森林结构研究的焦点[2]。准确提取森林空间结构信息,在空间结构分析的基础上制定合理的经营措施,改善和优化森林生态系统,这种森林空间结构化经营理论将成为未来森林可持续经营研究的一个重要方向[3-4]。
森林采伐是实现森林经营收获的一种手段,同时合理的择伐是调整林分空间结构、提升森林生态功能的重要手段。天然云冷杉林是长白山典型的地带性植被,也是当地主要的用材林和风景林,如何通过择伐的方式,既收获大量木材的同时,又能优化林分空间结构,实现天然林可持续经营是当前亟需解决的问题。因此分析过伐林的各种群生长过程和林分结构,了解伐后林分空间结构的动态变化特点至关重要。以往关于采伐方式对天然林的影响已有较多研究,主要集中在采伐对林分或树木生长量和生长过程的影响[5-6]、采伐后天然林更新的空间分布格局[7-10]、伐后生物多样性的评估及变化动态[11-12]等方面,而对于不同择伐类型天然林优势种群空间结构对比分析的研究[13],尤其是关于长白山天然云冷杉林的报道很少。
本研究利用描述空间点格局分布的O-ring统计方法,通过对长白山金沟岭林场的天然云冷杉林择伐固定样地的连续调查,就林分枯损、进界、主要种群空间分布格局和种间关联性等方面,研究其在3种不同择伐强度(弱度20%、中度30%、强度40%)干扰下的林分空间分布格局,比较不同择伐强度对林分空间格局的影响,为制定合理的择伐优化经营方案和恢复顶级天然云冷杉群落提供依据。
1 研究区概况
研究区域位于吉林省汪清县境内东部金沟岭林 场 (130°5′~ 130°20′E,43°17′~ 43°25′N), 属吉林省东部山区长白山老爷岭山脉雪岭支脉。林场地貌属低山丘陵地带,海拔为550~1 100 m,阳坡较陡阴坡平缓,坡度10°~25°。该区气候属于季风性气候,年平均气温4 ℃左右,年平均降水量600~700 mm,雨量主要集中在7月份。土壤属于低山灰化土灰棕壤区,一般为黏壤土类,土壤湿润、松散,根系多分布于土厚40 cm左右。
林场经营总面积为16 286 hm2,森林覆盖率为98%。该区的典型森林植被是以云杉Picea jezoensis和 冷 杉Abies nephrolepis为 建群种的天然暗针叶林,其他乔木树种主要有红松Pinus koraiensis、 枫 桦Betula costata、 白 桦Betula platyphylla、 紫 椴Tilia amurensis、 色 木槭Acer mono、 水 曲 柳Fraxinus mandcshurica、山杨Populus davidiana、杂木(青楷槭Acer tegmentosum、花楷槭Acer ukurunduense)等。
2 研究方法
2.1 样地设置及数据调查
本研究采用的4块云冷杉林样地均于1986年设置,样地是将0.64 hm2的样地均分为4块40 m×40 m小样地,并分别设置为弱度(20%)、中度(30%)、强度(40%)3种择伐类型和1块未择伐对照的固定样地,样地立地条件基本一致;每块样地初始择伐当年,调查样地的林分因子,包括林分起源、海拔、坡度和坡向等。将小样地分割成5 m×5 m的调查单元,以每个调查单元的一条边为x轴,以另一条垂直边作为y轴,记录乔木胸径≥5 cm的每一株林木的坐标值,同时每木检尺,记录其树种、胸径和树高。以后基本每隔两年复测1次,并记录进界木、枯死木以及更新等。本文所用数据为2008年(伐后第22年)复测数据,表1为不同择伐类型刚择伐后与伐后22年(2008年)的林分主要特征。
2.2 空间点格局分析
对种群空间格局的研究方法大致可分为样方法、角尺度法和距离法[14]。其中距离法中的点格局分析法是把每个植物个体都视为二维空间平面中具有坐标值的点,这样样地中的所有植物个体视为由一系列点组成的集合,构成点图;点格局分析法就是以点图为基础,描述不同尺度下种群空间格局及种间关联性的分析方法[15]。目前,二阶Ripley’s K函数广泛应用于种群生态学中[16-23],O-ring统计是在Ripley’s K函数和Mark相关函数的基础上发展的一类邻体密度函数[24],通过计算点在某一距离的发生的频率来分析其空间格局。该方法用圆环替代了Ripley’s K函数中使用的圆圈,计算与中心点某一距离的环内目标点的平均数目和分布状态,有效地消除了Ripley’s K函数随着尺度增大,存在着统计小尺度的累积效应,使格局分析复杂化的缺点[25]。O-ring统计研究包括单一种群空间格局的单变量O-ring统计和用于研究两个种群之间空间关联性的双变量O-ring统计。双变量O-ring的统计值[26]为:
式(1)中:n1为格局1的点的数目;[]表示以格局1第i个点为圆心、半径为r、宽为w的圆环;Points2[X]计算了区域X内格局2的点数目;Area[X]为区域X的面积。单变量统计是通过假设2种格局的点相同来进行计算的。
在本研究中,单变量O-ring统计用于分析不同择伐类型的天然云冷杉林主要树种的分布格局,双变量O-ring统计用于分析主要树种间的空间关联性。在分析时,要认真选择零假设,否则就会引起空间格局的误判。对于单变量分析,应先通过样地调查得到的每木定位图判断研究物种是否表现为明显的聚集性分布,如果不呈现明显的聚集性分布,采用完全空间随机零假设(complete spatial randomness,CSR);否则,则选用异质性泊松过程(heterogeneous poisson process,Poisson)。对于双变量分析,比较样地内3种建群树种之间的空间关联性,采用完全空间随机零假设。
样地的各树种分布图通过R软件(2.15.0版)绘制,O-ring统计分析使用基于栅格分析的生态学软件Programit(2008版)完成,空间尺度采用0~20 m,步长为1 m,Monte Carlo 循环模拟99次,将模拟结果中第5个最大、最小模拟值之间的数值围成99%的置信区间[27]。
3 结果与分析
3.1 不同择伐样地伐后的林分特征
调查统计资料(见表1和表2)表明,样地的主要乔木树种均为当地的顶级树种,树种组成中种类和各树种所占比例基本一致,云杉、冷杉2种顶级针叶树种均处于绝对优势地位,2个树种的蓄积总和占林分总蓄积的80%以上,其中冷杉占50%,云杉占20%,其他树种红松占10%,阔叶树以椴树和枫桦为主。不同样地中蓄积生长率随着择伐强度的增加而增大,3块择伐样地增幅明显高于对照样地。择伐22年后除强度择伐样地的林木株数增加外,其余样地都减小,其中对照和中度择伐样地的株数减小幅度较大,弱度择伐不明显。对照样地中的阔叶树的平均胸径相对较高,弱度择伐样地中各主要针叶树种的平均胸径(冷杉22.7 cm、云杉22.5 cm、红松24.2 cm)明显高于其他样地,中度择伐样地由于样地内小树较多,导致各树种和林分平均胸径较低。
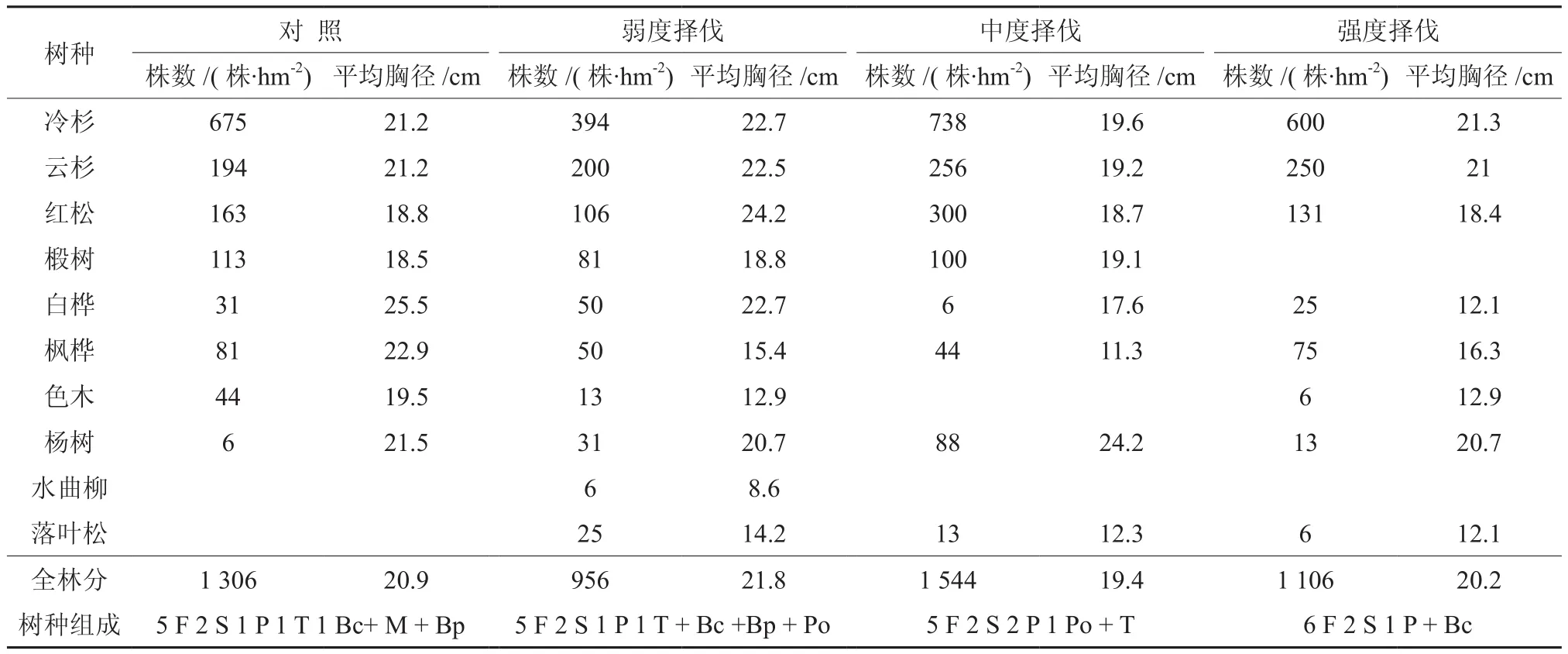
表2 不同择伐类型伐后22年乔木层主要树种基本概况†Table 2 Basic Situations of dominant species of 22-year-old tree layer with different selection cutting
3.2 不同择伐类型主要树种的空间分布格局
由对照样地和3块择伐样地的各树种分布图(见图1)可知:采伐并未明显改变林分整体分布状态,但在某些局部范围内不同树种和不同径级的林木密度差异较大,其密度随空间位置变化较大。
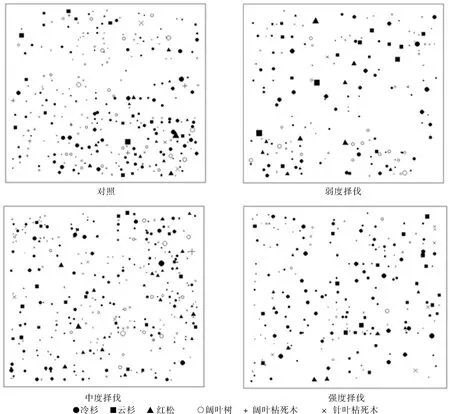
图1 不同择伐类型样地的树种空间分布Fig. 1 Spatial distribution of tree species in different selection cutting plots

表3 不同择伐类型主要树种的空间分布格局†Table 3 Spatial distribution patterns of dominant tree species in different selection cutting types
22年后不同择伐类型天然云冷杉林的优势种群不同尺度上的空间分布格局见表3。林分所有树木在1 m尺度上都表现为均匀分布,大于5 m尺度时分别出现不同尺度的聚集分布,其中39号和37号样地表现明显,分别在10~11 m和 15~16 m尺度内呈现显著的聚集分布。冷杉分别在39号样地5、10、15 m,37号样地的16~18 m尺度上呈现聚集分布,38、40号样地主要表现为随机分布,在个别尺度上表现为均匀分布。云杉在3块择伐样地的5 m和8 m尺度上分别呈聚集分布,并且在较大尺度上出现随机分布和均匀分布。红松的空间分布格局在不同尺度呈现为聚集分布,7 m尺度上择伐样地的聚集现象表现一致。
不同优势树种在各择伐类型样地中分布的比较表明:在所有样地中林分整体和各个主要树种均以随机分布为主导地位,部分尺度呈现聚集分布,较大尺度表现为随机分布和均匀分布;冷杉、云杉及林分在未择伐样地和强度择伐样地呈现的聚集分布比其他两种择伐类型明显,而弱度和中度择伐样地在不同尺度上呈现较多的均匀分布;红松在所有样地中均表现为较明显的聚集分布。
3.3 不同择伐类型优势种群的种间关联性
不同择伐强度样地主要树种相互的空间关联分析(见表4)表明:冷杉和云杉在对照样地和37号样地的1~2 m、38号样地的3~4 m尺度范围内分别呈现显著的负相关,40号样地在4 m尺度以及37号样地的15 m尺度表现为正相关。冷杉和红松在对照样地的3~4 m,40号样地的3、7、15 m尺度上呈显著正相关,38号样地18~19 m尺度内呈现显著负相关,40号样地在1、16 m尺度呈正相关。云杉和红松在4块样地不同尺度上主要表现为不相关,部分表现为负相关,38号样地在7、14~15 m尺度内负相关显著。
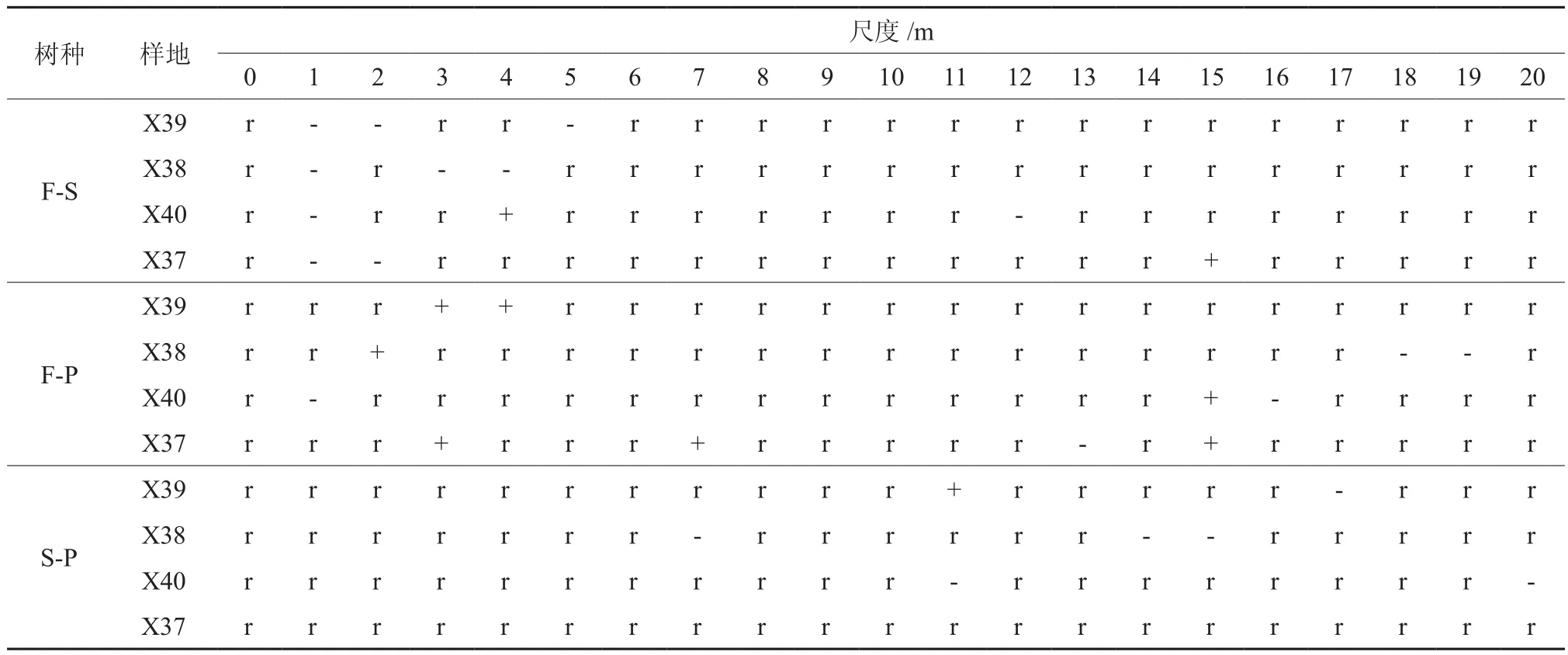
表4 不同择伐类型主要树种之间的空间关联性†Table 4 Spatial associations of dominant tree species in different selection cutting types
通过比较可知:不同树种之间的关联性以不相关为主,云杉和其他两种主要树种负相关性明显,而冷杉和红松之间呈现不同尺度上的正相关性。冷杉和云杉在小于5 m的尺度范围内均表现为强烈的负相关性,冷杉和红松种间呈现较多的正相关性,云杉和红松种间的关联性主要表现为不相关,不同尺度上出现负相关现象,弱度择伐样地的负相关性比其他样地表现显著。
4 结论与讨论
本研究的样地中冷杉、云杉和红松在密度和断面积中的比重均占主要地位,林分山杨、白桦等先锋树种所占比例较少,说明了调查地区正处于顶级群落。择伐没有改变林分内各树种所占的比例,但促进了天然云冷杉林林分的生长,择伐样地蓄积量的生长均大于对照,弱度择伐样地的冷杉、云杉和红松的平均直径明显高于其他,并且株数变化相对稳定。这与胡云云等[5]的结论一致。伐后22年间,相对择伐样地,对照样地表现为较高的枯损,这是因为进行采伐作业时优先选择处于劣势生态位的树木,一次性地伐去了这些树木。
不同择伐强度下的种群空间分布格局存在差异,这与种群进界个体的产生、生长劣势个体的死亡以及人为干扰有关。虽然在不同强度采伐下的林分以及优势种群仍然呈现以随机分布为主体的空间分布格局,但天然云冷杉林通过择伐经营,形成光照强弱、林隙效应以及土壤营养条件不均匀等空间异质性,造成了林分内树木的进界与枯死,导致不同尺度上的空间格局产生差异。红松在3个择伐样地内聚集现象较明显,这源自于其本身弱阳性的生物学特性和对林隙的响应,均匀分布的林隙和上层林冠的疏开为红松幼树生长提供了空间和机会[28]。林分及各优势树种随着择伐强度的增加聚集现象呈现先减小后增大的变化,说明人为干扰会改善林分中的聚集分布状态,弱度和中度择伐强度的林分趋于随机和轻微均匀分布,但当择伐超过一定强度,聚集性反而变大。这主要是由于择伐强度过大创造了许多大的林窗,地温增加,枯落物分解速度加快,某些喜光的树种加速生长导致林分整体聚集性增加。弱度择伐的林分及各建群优势种的径级、龄级相要比其他样地大,这些大树对林分小环境内有限的光、养分和水分等资源的需求就相对较多,相互之间对资源的竞争就相对强烈,从而形成自然稀疏,使个体数量递减,一定程度上解释了样地中种群分布聚集现象相对较少的现象,而且最终将表现为均匀或随机分布[29-31]。
空间关联性是对植物种群在空间分布上的相互关系的静态描述,这种关系涵括了空间位置关系和个体间功能关系[32]。不同择伐强度下各优势树种之间大多无相关性,不同尺度上的种间关联性表现有差异。在所有不同尺度上,冷杉、云杉和红松3种优势树种两两间的关联性,择伐样地与对照样地在不同尺度上表现的正相关与负相关现象有差异,而且弱度择伐相对其他择伐类型的负相关性现象多,这说明择伐强度改变了林分内各树种之间的竞争状态。冷杉和云杉在小尺度上表现出明显的负相关性,这是因为生态习性相近的种,种间竞争势必激烈,而冷杉作为优势度最大的树种,处于该林分中最适的生态位,拥有最多的空间和养料资源,邻近的云杉在小尺度上受到冷杉的排斥。弱度择伐样地中树木个体径阶较大,幼小个体对养分资源的竞争力微弱,但所有个体对土壤养分和空间资源的要求相同,由于单位面积上资源有限无法满足它们的需求,种间和种内必然产生强烈的排斥和竞争,呈现出的负相关相对较多。但强度择伐造成林分内单位面积上有充足的养分资源,能够满足各个树种的保留木和幼树的生长,导致各树种间聚集现象增多,负相关现象减少。
通过不同尺度的林木点格局分析,可以发现林分空间分布的特征点,这些特征点在实施经营措施时具有指导意义,如聚集分布的区域应是采伐的关注的地方,稀疏的区域应减少采伐,避免更大的稀疏。林分的整体均匀分布,有利于植物利用光照,调节林内小气候,促进林木及林下林下植物生长;同时使地表连续覆盖,符合近自然林业的要求。林分经营管理时应选择合适的采伐强度和采伐策略,本研究对主要由云杉、冷杉等阴性树种组成的天然云冷杉林进行优势种群空间分布格局和空间关联性进行分析,证明采伐强度20%(弱度择伐)更能促进天然云冷杉林群落结构优化,同时也利于云冷杉的生长。
[1] 汤孟平. 森林空间经营理论与实践[M]. 北京: 中国林业出版社, 2007.
[2] Moeur M. Characterizing spatial patterns of trees using stemmapped data[J]. For. Sci., 1993, 4(39):756-775.
[3] 惠刚盈,胡艳波,赵中华. 再论“结构化森林经营”[J]. 世界林业研究, 2009, 22(1): 14-19.
[4] 惠刚盈,Klaus von Gadow,胡艳波,等. 结构化森林经营[M]. 北京: 中国林业出版社, 2007.
[5] 胡云云,闵志强,高 延,等. 择伐对天然云冷杉林林分生长和结构的影响[J]. 林业科学, 2011, 47(2): 15-24.
[6] 蒋子涵,金光泽. 择伐对阔叶红松林主要树种径向与纵向生长的影响[J]. 生态学报,2010, 30(21): 5843-5852.
[7] 郑丽凤,周新年,罗积长,等. 择伐强度对天然针阔混交林更新格局的影响[J]. 福建林学院学报,2008, 28(4): 310-313.
[8] 韩景军,肖文发,罗菊春. 不同采伐方式对云冷杉林更新与生境的影响[J]. 林业科学,2000,36(S1): 90-96.
[9] 乌吉斯古楞,王俊峰,郑小贤,等. 金沟岭林场过伐林更新幼苗空间结构分析[J]. 中南林业科技大学学报,2009,29(4):21-25.
[10] 刘 云,侯世全,李明辉,等. 两种不同干扰方式下的天山云杉更新格局[J]. 北京林业大学学报,2005, 27(1): 47-50.
[11] 金永焕,李敦求,姜好相,等. 择伐干扰后长白山区天然次生林物种多样性的变化[J]. 吉林农业大学学报,2006,28(1):35-39.
[12] 邱仁辉,陈 涵. 择伐作业对常绿阔叶林乔木层树种结构及物种多样性的影响[J].中国生态农业学报,2005,13(3):158-161.
[13] 郑丽凤,周新年. 择伐强度对中亚热带天然针阔混交林林分空间结构的影响[J]. 武汉植物学研究,2009, 27(5): 515-521.
[14] 惠刚盈,李 丽,赵中华,等. 林木空间分布格局分析方法[J].生态学报,2007, 27(11): 4717-4728.
[15] 张金屯,孟东平. 芦芽山华北落叶松林不同龄级立木的点格局分析[J]. 生态学报,2004,24(1): 35-40.
[16] Lingua E, Cherubini P, Motta R, et al. Spatial structure along an altitudinal gradient in the Italian central Alps suggests competition and facilitation among coniferous species[J]. Journal of vegetation science, 2008(3):425-435.
[17] Rozas V. Structural heterogeneity and tree spatial patterns in an old-growth deciduous lowland forest in Cantabria, northern Spain[J]. Plant Ecology, 2006(185):57-72.
[18] 龚直文,顾 丽,亢新刚,等. 长白山森林次生演替过程中林木空间格局研究[J]. 北京林业大学学报,2010,32(2): 92-99.
[19] 李 立,陈建华,任海保,等. 古田山常绿阔叶林优势树种甜槠和木荷的空间格局分析[J]. 植物生态学报,2010, 34(3):241-252.
[20] 汤孟平,周国模,施拥军,等. 天目山常绿阔叶林优势种群及其空间分布格局[J]. 植物生态学报,2006, 30(5): 743-752.
[21] 王 蕾,张春雨,赵秀海. 长白山阔叶红松林的空间分布格局[J]. 林业科学,2009,45(5): 54-59.
[22] 张 赟,张春雨,赵秀海,等. 长白山次生林乔木树种空间分布格局[J]. 生态学杂志,2008,27(10): 1639-1646.
[23] Salas C, LeMay V. Spatial patterns in an old-growth Nothofagus obliqua forest in southcentral Chile[J]. For. Ecol. Manag.,2006(231): 38-46.
[24] Lawes M J, Griff i ths M E, Midgley J J, et al. Tree spacing and area of competitive influence do not scale with tree size in an African rain forest[J]. Journal of Vegetation Science, 2008, 19:729-738.
[25] Wiegand T, Moloney K A. Rings, circles, and null-models for point pattern analysis in ecology[J]. OIKOS, 2004(104):209-229.
[26] 张金屯. 数量生态学[M]. 北京: 科学出版社, 2004.
[27] Dietrich S, Helga S. Fractals, random shapes and point fields:Methods of geometrical statistics[M]. New York: John Wiley &Sons, 1994.
[28] 黄新峰,亢新刚. 我国天然云冷杉针阔混交林更新研究进展[J]. 世界林业研究. 2004,17(5): 34-38.
[29] 闫海冰,韩有志,杨秀清,等. 华北山地典型天然次生林群落的树种空间分布格局及其关联性[J]. 生态学报,2010, 30(9):2311-2321.
[30] 周建云,李 荣,张文辉,等. 不同间伐强度下辽东栎种群结构特征与空间分布格局[J]. 林业科学,2012,48(4):149-155.
[31] 张笑菁,赵秀海,康峰峰,等. 太岳山油松天然林林木的空间格局[J]. 生态学报, 2010,30(18): 4821-4827.
[32] 王 磊,孙启武,郝朝运,等. 皖南山区南方红豆杉种群不同龄级立木的点格局分析[J]. 应用生态学报,2010,21(2):272-278.
Effects of selective cutting intensity on spatial distribution pattern of natural spruce-f i r forests
SHEN Lin, YANG Hua, KANG Xin-gang, YUE Gang
(Key Lab. for Silviculture and Conservation of Ministry of Education, Beijing Forestry University, Beijing 100083, China)
The research was done in order to explore the effects of different selective cutting disturbance of different intensities on the spatial patterns of natural spruce-f i r stands and provide evidences for reasonable management forest. The spatial patterns of dominant species and the spatial correlations among different species were analyzed by using one point pattern analysis method, which is O-ring statistics. The experimental stands were harvested by selective cutting of different intensities (low intensity of 20%, medium intensity of 30%, high intensity of 40%) and a un-cutting plots. The results show that the spatial pattern of either the forest as a whole or the dominant species took on a random distribution, that of the forest with strength selection cutting showed a aggregated distribution, while in random and uniform distribution with low and medium selective cutting. There existed a phenomenon that the negative correlations of dominant species with low intensity elective cutting were more than other cutting type’s. The low intensity selective cutting may effectively enhance the stability of forest community spatial structure and make the communities spatial structure more superior.
natural spruce-f i r stand; selective cutting intensity; spatial distribution pattern; O-ring statistics
S752.2
A
1673-923X(2013)01-0068-07
2012-10-10
林业公益性行业科研项目(200804027)
沈 林(1988-),男,天津人,硕士研究生,主要从事森林可持续经营研究;E-mail:33235213@qq.com
杨 华(1964-),女,北京人,副教授,主要从事资源与环境信息系统和森林资源监测技术研究;
E-mail:huayang8747@163.com
[本文编校:谢荣秀]
猜你喜欢
贵州畜牧兽医(2022年3期)2022-06-28
小学教学参考(语文)(2022年3期)2022-05-26
现代园艺(2021年23期)2021-12-01
新农业(2020年18期)2021-01-07
花卉(2020年24期)2021-01-04
云南医药(2020年5期)2020-10-27
防护林科技(2020年6期)2020-08-12
绿色科技(2019年6期)2019-04-12
绿色科技(2019年6期)2019-04-12
中国医药指南(2017年3期)2017-11-13
