
山丘区钉螺大小特征与水分依赖性
2012-11-28 09:35何亚平蒋俊明费世民陈秀明周金星张旭东彭镇华
四川林业科技 2012年1期
何亚平,蒋俊明,费世民**,陈秀明,周金星,张旭东,彭镇华
(1.四川省林业科学研究院,四川成都 610081;2.中国林业科学研究院,北京 10093)
钉螺是我国大陆上古老的动物,是血吸虫病流行的中间宿主,是血防中采取措施的对象。钉螺壳形态是钉螺分类学的重要指标,也是地理亚株划分的重要依据。周艺彪等(2006)将21个钉螺居群用螺壳形态数量聚类分析为3类;周晓农等(1997)对34个居群钉螺进行数值分类,结果表明壳体大小、壳形(光壳和肋壳)和壳厚是描述钉螺螺壳的主要特征。实际上这种螺壳大小特征可能也与环境因子紧密相关,即壳体类型、大小受到环境因子的调节。我国钉螺孽生数量、分布特性与环境因子关系研究很多(何亚平等,2006;许发森等,1999;蒋俊明等,2006;张旭东等,1998;项艳等,1999;吴立勋等,2004;张旭东等,1999;吴刚等,1999;王家生等,2007;郭玉红等,2011;董毅等,2009;梅勇等,2010;王万贤等,2009;陈柳燕等,2002;马安宁等,1999;汪奇志等,2001;熊晓娇等,2006;张家来等,1996;张旭东等,2007;朱中亮,1992),壳形,如光壳或肋壳就被认为与自然环境长期适应有关(胡相,1993)。关于钉螺大小与环境因子关系研究还很少涉及,限制了对钉螺在不同环境条件下生态适应性的探讨。
尽管数量与分布受到诸多因子的影响,而钉螺自身重量和壳体大小可能更大程度上与地理环境有关,并呈现明显的居群间分异与聚集性特点(周晓农等,1977;周艺彪等,2006)。温度对钉螺发育影响极为明显,在各温度下初孵幼螺随着生长发育,螺壳的长宽比也随之加大(张垒等,2008)。影响钉螺数量、分布的生物与非生物环境因子中都与土壤水分或地下水位有着密切关系(项艳等,1999;吴立勋等,2004)。然而其孽生地理环境或微观立地都存在水分与热量匹配,可能温度与水分的适宜组合塑造了钉螺的壳体长、宽、体积与重量等大小指标。
山丘型地区钉螺孽生的立地条件较为复杂,其生境因子组合异质性相对较强。山丘区孽生钉螺大小与壳体特性仅局限于中国疫区钉螺形态性状的聚类分析(周晓农等,1977;周艺彪等,2006),斜高和角度的地区性差异(王经邦,刘寿喜,1965a,1965b),对钉螺壳体长宽、鲜重和体积等大小指标与水热关系存在宏观区域与微观土地利用类型或生境两种尺度。
本文在山丘型钉螺分布区仁寿县研究了钉螺壳体长宽、鲜重与体积在微观生境条件下的现状,并借助发表数据分析了区域壳体长宽与降水量、温度的关系,研究探讨钉螺大小与水热因子的关系。
1 材料与方法
1.1 示范区自然地理概况
仁寿县属低山丘陵地貌类型,地势西北高、东南低,二峨山(龙泉山脉)将县境分成东南、西北两大部分。示范区位于二峨山东南部的丘陵区,因切割程度的差异,可分为浅丘和深丘,丘陵区占全县幅员面积的70.14%。仁寿属亚热带季风性湿润气候,冷热四季分明,冬无严寒,夏无酷暑,春多干旱,秋雨绵绵的特点,累年平均气温17.4℃,全年气温以7月最高,为26.5℃,1月最低,为6.8℃,全年极端最低温为-3.8℃,总正积温6 360℃,≥10℃的积温为5 532.8℃;全年无霜期310 d左右,最长356 d,最短282 d。历年平均日照1 196.6 h,年日照率27%,太阳总辐射89 kcal·cm-2,日照时数和总辐射为全国低值区。
仁寿复杂的土壤母质,紫色砂页岩占全县土壤母岩的86.9%,第四系黄色老冲积和新冲积,洪积物仅占5.7%,三迭系和自流井群占7.4%。二峨山东南土壤有紫色土类大土泥、黄沙土、冷沙黄泥土等,微酸至弱碱,土层较厚。由于地势高差较小,森林的垂直分布梯度不明显,地带性植被为常绿阔叶林。由于垦殖指数高,原生植被已全部被人工植被所替代,主要的林分类型为柏木林、马尾松林、马尾松-油茶混交林、竹林及四旁树,但按优势建群种分为马尾松和柏木两个片区,马尾松主要分布于山地黄壤和老冲积黄壤地区,柏木林分布丘陵的钙质紫色土地区,以慈竹为主的竹林主要分布于四旁,桉树、桤木、杨树等速生树种主要栽植于水分条件较好的田边地坎,溪沟河流两岸。
兴林抑螺小流域示范区建设在仁寿县洪峰乡新建3队。该示范区建设于2007年,该村血吸虫病解放前与解放后极其严重,是仁寿县的重灾区,由和尚沟、茶子林沟、青冈林沟、猫儿沟组成。主要森林斑块为柏木林、马尾松林、慈竹林、桉树林,水田集中在沟底与二台丘顶平台,向上坡面延伸有少量旱地。示范区建设前旱地较多,且水田本身田埂桑树、构树栽植相对较为普遍,只是经营性较差,处于荒芜状态。钉螺主要孽生于干渠、路渠、河溪、田渠、屋旁、旱地水沟、岩石洼地、荒芜水田、荒芜鱼塘等地带。
1.2 调查方法
2011年10月份,收集该兴林抑螺示范区内不同环境条件下钉螺,在50 cm×50 cm样方内收集所有钉螺,用铝盒带回,并记录生境特征。室内进行钉螺计数与鲜重测量,并选择典型钉螺进行长度、宽度测定。
查阅文献(周艺彪等,2006),对中国疫区不同地区气候资料进行网上查阅,并摘录钉螺长度、宽度数据,将本次仁寿县钉螺长度、宽度数据补充于上述数据中。计算单位降水量的温度值,该值能反映相对的干燥程度。数据如表2所示。
钉螺鲜重测定方法:计量钉螺数量、用铝盒盖装载放在电子天平上测定,记录重量,并将铝盒盖再次单独称量,二者之差为钉螺鲜重,并除以数量,求得单个钉螺鲜重。钉螺长度测量以螺口至顶部最大垂直距离为准,宽度为第一旋直径,测量工具为游标卡尺,单位为mm。数据如表1所示。

表1 仁寿新建3队猫儿沟小流域农林复合系统内钉螺大小与生境状况
钉螺体积计算以锥体体积公式来计算,即V=长度×宽度/3。
数据分析方法采用回归分析法,对钉螺大小与降水量、温度,以及钉螺大小自身参数进行散点作图,并拟合相关曲线。
2 结果与分析
2.1 仁寿钉螺大小特点
仁寿新建3队猫儿沟小流域农林复合系统内钉螺大小如表1所示。计算表明,钉螺鲜重为0.0162 g,浮动在0.0065 g~0.0219 g之间,长度为5.156 mm(3.6 mm~6 mm),宽度为2.668 mm(2.2 mm~3 mm),体积为4.5894 mm3(3.12 mm3~5.6 mm3)。钉螺大小特征在样地之间变异很大,变异系数分别为 4.038(鲜重),7.908(长度),16.675(宽度),6.899(体积),表明钉螺宽度变异最为明显,其次为长度和体积,变异最小的为鲜重。
钉螺长度级划分频度如图1所示。长度2 mm~3 mm钉螺占到0.361%(1个),长度3 mm~4 mm钉螺占到10.4693%(29个),长度4 mm~5 mm钉螺占到24.1877%(67个),长度为5 mm~6 mm钉螺占到43.6823%(121个),长度5 mm~6 mm钉螺占到21.2996%(59个)。表明,仁寿钉螺长度主要为4 mm~7 mm之间,占到80%左右,钉螺长度以5 mm~6 mm为主体,超过40%,而4 mm以下钉螺相对较少,总体占到11%。
钉螺宽度在1 mm~2 mm之间比例很低,仅有0.7194%,宽度2 mm~2.5 mm钉螺占到16.9065%(47个),2.5 mm~3 mm宽度钉螺比例为71.9424%(71.9424%),3 mm~3.5 mm宽度钉螺比例为10.4317%(10.4317%)。表明,仁寿钉螺宽度在2 mm~3.5 mm之间,而以2.5 mm~3 mm宽度钉螺为主体。

图1 钉螺长度与宽度的频度分布
2.2 钉螺大小参数间相关性
钉螺长度与宽度,钉螺鲜重与长宽之间散点图如图2、图3所示。钉螺长度与宽度之间散点图构成显著的直线关系(r=0.278,p=0.000),符合直线方程:y=0.0764x+2.2733,R2=0.0715。钉螺长度与宽度之间这种直线关系斜率相对较低,仅有0.0764,但从统计上说钉螺宽度随着长度增加而增加,即钉螺长度发育越大而宽度相对较大。但从散点图上宽度幅度来看,钉螺宽度范围相对较小,而长度范围相对较大,又暗示长度变化时宽度变化幅度或速度并不大,尽管统计检验上是显著的。从大小形态上分析,钉螺宽度相对稳定,而长度变大呈现较小的宽度变化,变化速率为0.0764;从发育上看,钉螺先发育宽度,后续螺旋逐渐发育,形成钉螺的长度发育过程,实际上小钉螺是高度不足。

图2 钉螺长度与宽度之间散点图
钉螺鲜重与长度、宽度、体积之间散点图如图3所示。统计分析,这些拟合直线都未构成显著关系(鲜重与长度:f=0.229,p=0.271;鲜重与宽度:r=0.341,p=0.095;鲜重与体积:r=0.204,p=0.327)。钉螺鲜重与长度、宽度、体积关系不明显,暗示鲜重与大小之间关系可能在群体上并不明显,群体平均缩小了个体间差异,大小参数与鲜重之间关系表现在居群内个体之间。在群体水平上,钉螺鲜重可能与环境有关。

图3 钉螺鲜重与长度、宽度、体积散点图
2.3 钉螺大小与环境关系
仁寿县兴林抑螺示范区不同钉螺大小与环境特点如表1所示。在一些旱地内侧坡坎地带,如样地12-5、12-6、13-19、12-9 钉螺鲜重为0.0065 g~0.0168 g,体积分别为 3.447 mm3~4.95 mm3,长度为4.2 mm~5.5 mm,宽度为2.22 mm~2.9 mm,而水稻田坡坎,如样地 13-1、13-2、13-4、13-3、13-8、13-7,钉螺鲜重为0.0156 g~0.0217 g,体积为4.247 mm3~5.6 mm3,长度为4.9 mm ~6 mm,宽度为2.6 mm~2.9 mm;水淹出水湿润生境,钉螺鲜重为0.0137 g~0.0166 g,体积 3.467 mm3~4.737 mm3,长度4 mm~5.1 mm,宽度为2.5 mm~2.9 mm。统计分析结果表明,湿润浸水生境、旱地生境与水稻田生境间钉螺长度差异极为明显(f=5.026,p=0.026),而鲜重、体积、宽度等差异不明显(f=0.726,p=0.504;f=2.725,p=0.106;f=0.004,p=0.996)。河岸、泉水出露、浸水田缘等土壤水分湿润,旱地在水分季节性潮湿,水田则潮湿相对稳定,钉螺长度差异相对更为明显,暗示钉螺大小与微观生境水分含量关系密切。钉螺长度发育与水分存在联系,在泉水、河水富集地带,钉螺往往长度较小,而在水分不足旱地生境中长度较为充足,而潮湿地带钉螺长度适宜,可能温度也是促进长度的因子。此外,鲜重可能与环境水分有关,群体上差异不明;宽度发育早,差异不明;长度因温度适宜而发育充分,相对在不同环境之间呈现了明显的差异性。
2.4 钉螺大小与区域降水、温度的关系
用中国疫区钉螺长度、宽度与降水量、温度数据(如表2所示)进行散点图和曲线拟合(如图4、图5所示)。其中温度与降水量比值反映了单位降水量背景下的温度,实际上蒸发状况,可反映实际干燥程度。实际上平均温度区域之间差异小,降水量差异相对较大,也是水网型、湖沼型与山丘型地区水热差异。
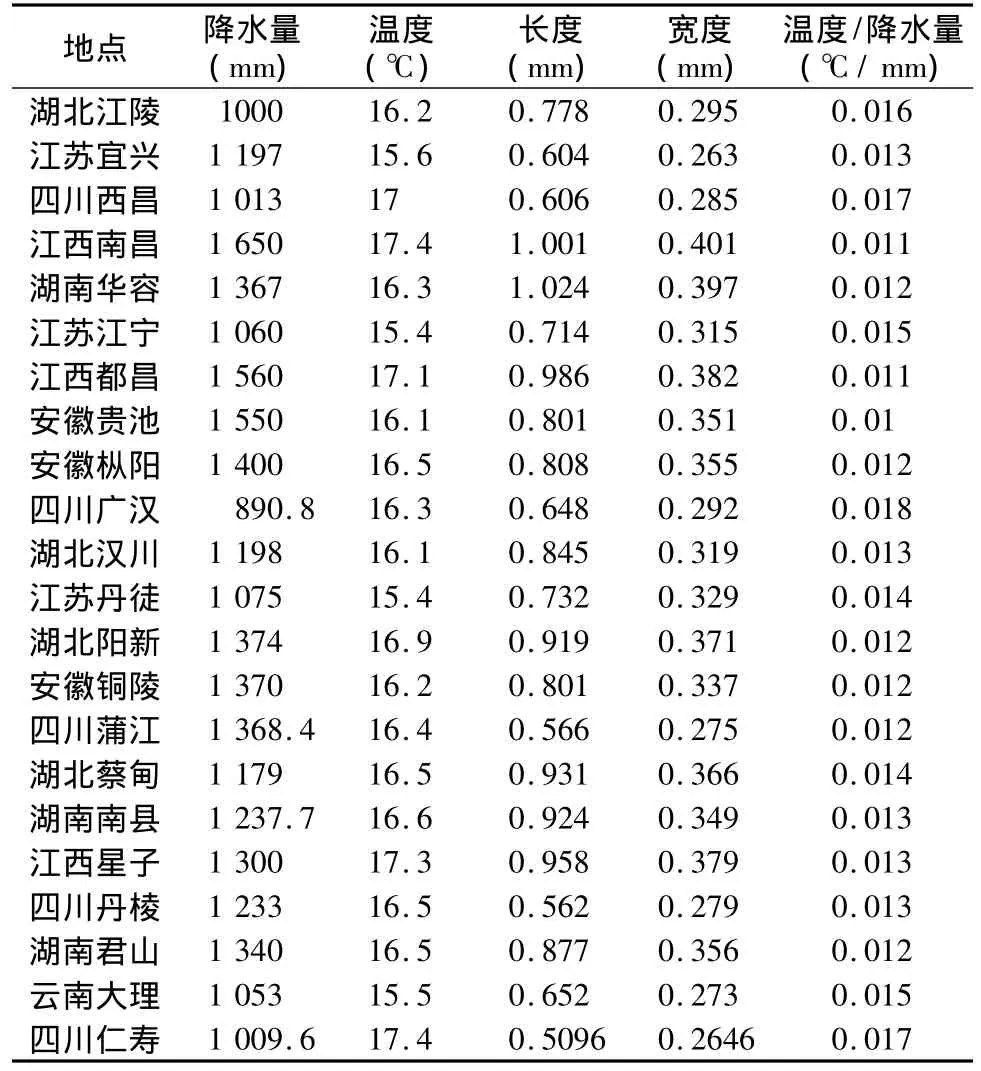
表2 中国典型疫区的降水量、温度(及温度/降水量比)与其孽生钉螺的长度、宽度数据[不同疫区钉螺长度、宽度数据源自于周艺彪等(2006),降水量、温度数据为Internet提供,仁寿钉螺长度、宽度为补充测定]
中国各疫区钉螺宽度、长度与降水量、温度之间散点图如图4所示。钉螺长度与降水量之间拟合曲线关系显著(r=0.596,p=0.003),长度与温度之间拟合曲线关系不明(r=0.288,p=0.193);钉螺宽度与降水量之间拟合曲线关系显著(r=0.691,p=0.000),与温度之间拟合曲线关系不明(r=0.364,p=0.096)(如图4所示)。温度降水量比例与钉螺长度之间拟合曲线关系显著(r=-0.563,p=0.006),与钉螺宽度之间拟合曲线关系显著(r=-0.624,p=0.002)(如图5所示)。表明,钉螺长度、宽度受到降水量影响较为明显,但不受温度的影响,但受到温度与降水量比值的影响,暗示在相对较为干燥时钉螺长度、宽度相对较小。这一点与水网湖沼地区钉螺较大而山丘区钉螺相对较小(周晓农,2004)的结论相一致,也是水热匹配所致,即在降水量低而温度相对较高地区钉螺相对较小。从图5还可知,钉螺长度随着温度降水量比递变速度较快,而宽度递变较慢,二者直线斜率明显不同(y=-41.923x+1.3461,R2=0.317;y=-13.164x+0.5053,R2=0.3895)。

图4 中国各疫区钉螺宽度、长度与降水量、温度之间散点图

图5 中国各疫区温度降水量比与钉螺长度、宽度散点图
3 讨论
钉螺孽生也是钉螺对适生环境的适应过程,也是不同生境下的生存适合度问题。钉螺数量分布明显与植被或林分特征(张旭东等,1998,1999;吴刚等,1999;董毅等,2009;郭玉红等,2011;王万贤等,2009;马安宁等,1999;熊晓娇等,2006;张家来等,1996)、土壤类型或组成(许发森等,1999;蒋俊明等,2006;项艳等,1999;吴立勋等,2004;张旭东等,2007;汪奇志等,2001;朱中亮,1992)、气象或温度(欧阳红等,1999;梅勇等,2010;陈柳燕等,2002;张垒等,2008;梁幼生等,1996)、人为活动或土地利用类型(何亚平等,2006;王家生等,2007)有关,在微观分布格局上,也表现为稻田坡坎下部明显较为密集,而中部稀少,上部零星或没有的现象。在小流域尺度上,钉螺表现出沿水系连续分布或离散分布的格局;在县级区域尺度上,山丘型钉螺往往表现出局部分布而整体上不均一的景观格局,山系、森林植被往往是钉螺分布的上限,且人为活动与钉螺孽生耦合性强,并部分地区表现为有螺无病现象(仁寿县血防志,1980)。这充分阐明,钉螺数量与分布跟植被、土壤、气象因素有关,并在中国长江流域呈现带状局限分布(朱中亮,1992)。这种影响数量分布的因子可能直接影响到钉螺大小,进而影响到钉螺的生存适合度,最终影响钉螺居群的发展动态。
山丘区钉螺明显较小,具有较小的高度和宽度,如西昌钉螺长 0.606 mm、宽0.285,广汉钉螺长0.648 mm、0.292 mm,蒲江钉螺长 0.566 mm、0.275 mm,丹棱钉螺长0.562 mm、0.279 mm,大理钉螺长0.652 mm、0.273 mm(周艺彪等,2006),仁寿钉螺长0.509 6 mm、0.264 6 mm(本文测定)。山丘区钉螺明显小于湖沼型和水网型,长度为0.562 mm~0.652 mm,宽度为0.275 mm~0.292 mm,而其他类型钉螺长度为0.604 mm~1.024 mm,宽度为0.203 mm~0.401 mm(如表2所示)。长江下游也有丘岗地带,山丘型在湖沼、水网向山丘、山岭延伸的地带也有钉螺分布,其本身也有山丘型钉螺,如江苏宜兴县,该处钉螺长度(0.604 mm)、宽度(0.263 mm)都明显较小。在微观分布上,调查结果表明,水稻田钉螺长度相对较大,旱地生境长度相对较小,过于湿润的水岸、泉水生境中钉螺长度也相对降低。但鲜重、体积、宽度则无明显差异,表明群体上这些特征并未受到生境异质性的调节,尽管还有钉螺种群自身大小与年龄结构的问题。仁寿县不同生境土壤水分、温度调查结果表明,沟系河滩水陆交错带土壤水分为28.08% ~30.48%、土壤温度为 20.5℃ ~20.73℃,旱地21.9% ~31.78%、17.86℃ ~19.5℃,水田32.5%、20.8℃(黄玲玲,2006)。水分差异导致了水稻田钉螺发育充足,具有较大的长度,而旱地钉螺宽度相对降低,尽管不显著,长度发育充足但总体还是短于水稻田钉螺,而湿润生境钉螺发育代间重叠明显,可能是大小结构异质性降低了总体的钉螺大小。3种生境水分差异体现在旱季,而在雨季相对差异也存在,即旱地相对干旱一些,而旱季则水稻田排水沟相对潮湿,而旱地则明显干燥,总体水分储量上应该是水流沟系大于稻田,而稻田大于旱地。钉螺长度这种差异表明,钉螺发育的微观水分含量决定了钉螺的长度大小,其大小具有水分依赖性。
实际上,钉螺发育也存在速率的问题,或者速率影响了壳体大小,即幼螺发育时高温高湿则直接快速形成壳体,而干旱或低温时新陈代谢慢,壳体形成慢,或者滞育了。调查中,发现很多滞育钉螺在水岸、稻田泉水出露地带呈现,即宽度达到山丘型钉螺的大小标准,而长度则明显不足。并且,这些地带钉螺交配产卵时间长,卵随时都在孵化,钉螺年龄结构复杂。随着春季温度的升高,钉螺孵化时间缩短,如13℃时开始孵化平均天数为35 d、全部孵化平均天数 50 d,16℃时 24 d、33 d,25℃时,15 d、20 d(陈柳燕等,2002)。利用有效积温公式得知,钉螺卵孵化的有效积温为193.55日度,发育起点温度为10.86℃;在各温度下初孵幼螺随着生长发育,螺壳的长宽比也随之加大(张垒等,2008)。钉螺发育的速度可能与3类生境的温度有关,水分充足时,地表温度相对较低,而水分相对限制时,地表温度相对较高。显然,基于钉螺发育时水分、温度的重要性,表现为水热匹配性,在一个因子不足时另一个因子相对充足时也能起到补偿性作用。部分生境钉螺温度较低,但水分相对较为充足,钉螺也能很好发育,但钉螺宽度、长度相对较小(如屋后积水地钉螺的发育);同理,在水分季节性明显时,温度相对充足则钉螺也能很好发育,如旱地生境,即高温加快钉螺发育的速度(细而高)。从生活史理论推论,在资源有限时,发育时间段而快则钉螺较为细高,而发育时间长时钉螺粗短(斜高小和角度大);在水热匹配较好时,即资源充足时,钉螺高度发育能建立在较大的宽度上,成螺又粗又高(斜高大和角度大)。钉螺发育呈现宽度先成而高度后成的特点(如图2所示,直线缓平,但检验显著),滞育钉螺次螺层叠加在一起,是没有良好伸张的结果,与幼螺发育时生境温度和水分供应有关。这样,就完全建立了钉螺大小形成的水分、热量匹配性模型,并在不同尺度上都可运用。山丘区水分和热量匹配上降水、热量普遍偏低,在山丘地形,积温相对更低,钉螺发育在秋冬交界时发生滞育很正常,钉螺都呈现矮小的状态,普遍小于水网和湖沼地区的钉螺(周艺彪等,2006;周晓农,2004)。
在中国长江流域整体疫区上考虑钉螺壳体大小发育问题,则明显呈现出降水量与钉螺长、宽线性相关性,但温度并不是影响成螺壳体大小的因子(如图4所示)。用单位降水量来计量水热匹配性时,则钉螺长度、宽度都与这一温度降水量比值存在显著的线性相关性,且长度随之变化迅速而宽度随之变化缓慢(如图5所示),表明水分和热量匹配性也影响了钉螺壳体长宽的大小。从表2数据可知,山丘区实际上存在降水量和温度二者匹配性的问题,如广汉、仁寿、西昌、大理温度降水量比例较高(0.0018、0.0017、0.0017与0.0015),丹棱、蒲江温度降水量比值较低(0.0013和0.0012),后者钉螺长度分别为 0.562 mm、0.566 mm,宽度为0.279 mm、0.275 mm;前者钉螺长度为0.648 mm、0.606 mm、0.652 mm,宽度差异相对较小(0.292 mm、0.285 mm、0.273 mm)。仁寿钉螺测定为深丘地区,其平坝钉螺可能与水热数量相适宜,可能相比之下,深丘钉螺大小相对偏低。由于水分因子对温度存在补偿作用,即在温度较低时,水分充足也能保证钉螺的持续发育;温度自身在疫区形成与钉螺大小塑造上与水分条件相结合才能发挥影响。单纯温度对钉螺的影响是发育速率的问题,或者生境选择的问题,良好的水热匹配才有较大的钉螺形成。这一点在表2和图5中反映较为明显,区域上钉螺差异大小是水热匹配共同塑造的,如湖沼型钉螺较大,水分充足而温度适宜,钉螺发育速度快,壳体较大,水网地区冬季水分较差,而热量充足,钉螺壳体相对稍小,而在山丘区则降水一般而温度偏高,钉螺发育时水分总体上呈现季节性限制,钉螺壳体相对偏小。环境梯度条件下钉螺大小与水分、热量匹配性模型如图6所示。
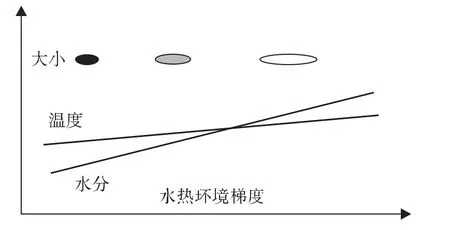
图6 环境梯度条件下钉螺大小与水分、热量匹配性模型
总体上看,水分供应越充足,钉螺壳体长宽越大;但在各降水量段内,温度与土壤水分匹配性也直接决定了钉螺壳体大小。区域上降水量梯度分异塑造了钉螺壳体大小,而局部水分配置格局直接塑造了钉螺壳体大小段内的变异性。区域整体上水分是温度与降水量二者共同决定的结果;微观上,钉螺发育的水热条件是土地利用类型与水分配置格局共同作用的结果,土表遮阴与否是微观上温度高低的影响因子。但水分极为充足时,温度较高时,季节内钉螺发育代内重叠极为明显。
钉螺是陆地上较为古老的物种,其分布位于水陆交错带(水系出露)和农林交错带(泉水出露),分布范围局限于第四系黄土层(朱中亮,1992),与这种土壤吸水松软而储水能力强易于穴居有关。钉螺物种具有自身生物学特性,并具有一定的个体大小变异性范围,尽管区域上大小差异显著而种下地理亚种间分化明显(周艺彪等,2006)。从分布上考虑,山丘区是钉螺分布的北部分界线,在江苏、安徽、湖北等地也有山丘区类型,其钉螺小,已经形成了独立的分化亚株,并在山丘区深丘地带,长度已经降低为湖沼和水网地区的一半。尽管宽度保持了相对稳定性,或低变异性,但有些钉螺宽度也明显高于山丘区宽度1 mm,发生交配上的不适性,呈现局域隔离种群状态。在山丘区钉螺种群内部,由于水分配置季节性分化明显,也使得部分生境下钉螺更为细小,如旱地生境。局域种群个体缩小是种群生境不适性增强的表现,这种不适性与生境的水热匹配有关。由于山丘区仁寿县林业血防安全工程体系建设相对较好,水岸、路渠、渠埂、人居等地血防林发达,整体上形成了不利于健壮钉螺发育的生境;从区域整体上水资源配置上考虑,较高的血防林比例增加了土壤水分耗用与降水截留,使得钉螺种群维持需要的地表水范围缩小,钉螺分布区萎缩而大小降低,呈现衰退状态。更为主要的是,这种血防林安全体系上水资源配置优化,使得依赖于地表水的血吸虫也无法完成生活史,加之改良的农业生产技术(水旱轮作),隔离了钉螺与血吸虫,而得二者都呈现种群衰退萎缩的趋势。这种衰退首先是分布范围缩小,密度降低,其次是大小变小。这一点在仁寿县钉螺密度调查中得到了很好的反映,尽管一些死角还存在高密度钉螺的分布,如路旁岩石洼地、田埂下泉眼处、干渠等。林业血防工程作用发挥首先表现为分布范围萎缩,其次为大小变小,本文就研究了仁寿县山丘区兴林抑螺示范区钉螺大小与生境水热异质性匹配的问题。
[1]周晓农,洪青标,孙乐平,等.中国钉螺螺壳的聚类分析[J].动物学杂志,1997,32(5):4 ~7.
[2]周艺彪,姜庆五,赵根明,等.中国大陆钉螺螺壳形态性状聚类分析[J].动物分类学报,2006,31(2):441 ~447.
[3]何亚平,费世民,蒋俊明,等.仁寿县钉螺数量和空间分布特点[J].湿地科学与管理,2006,2(4):40 ~43.
[4]许发森,钱晓洪,文松,等.安宁河流域植物、土壤特征与钉螺分布的关系[J].四川动物,1999,18(2):62 ~63.
[5]蒋俊明,何亚平,费世民,等.山丘型地区钉螺孽生数量与植被和土壤环境因子的关系[J].湿地科学与管理,2006,2(4):33~39.
[6]张旭东,吴刚,彭镇华.滩地林业生态工程与钉螺孳生关系的研究[J].应用生态学报,1998,9(5):468 ~470.
[7]项艳,彭镇华,张旭东.滩地地下水位对钉螺分布密度和I-72杨生长量的影响[J].安徽农业大学学报,1999,26(2):208~211.
[8]吴立勋,汤玉喜,吴敏,等.滩地钉螺种群消长与杨树人工林关系的研究[J].湖南林业科技,2004,31(6):5 ~9.
[9]张旭东,杨晓春,彭镇华.钉螺分布与滩地环境因子的关系[J].生态学报,1999,19(2):265 ~269.
[10]吴刚,苏瑞平,张旭东.长江中下游滩地植被与钉螺孳生关系的研究[J].生态学报,1999,19(1):118 ~121.
[11]欧阳红,欧阳芳芳,冯慧.气象要素与血吸虫钉螺繁衍内在规律[C].第26届中国气象学会年会气候环境变化与人体健康分会场论文集.2009.
[12]王家生,卢金友,魏国远,等.环境变化对钉螺扩散影响规律研究[J].长江科学,2007,24(3):16~19.
[13]郭玉红,郎南军,杨文灿,等.云南高原山地林分郁闭度与钉螺密度关系的研究[J].湿地科学与管理,2011,7(1):14~15.
[14]董毅,董兴齐,冯锡光.山丘地区植被指数与钉螺分布的关系[J].中国血吸虫病防治杂志,2009,21(2):98 ~101.
[15]梅勇,唐云辉,杨世琦,等.巫山大昌消落带钉螺可能孽生地气候生态环境分析[J].中国农业气象,2010,31(4):591~595.
[16]王万贤,杨毅,程炯,等.长江中下游江滩防洪林对钉螺分布密度影响[J].水生生物学报,2009,33(2):310 ~315.
[17]陈柳燕,徐兴建,杨先祥,等.三峡建坝后江汉平原土壤含水量及气温对钉螺生态的影响[J].中国血吸虫病防治杂志,2002,14(4):258 ~260.
[18]马安宁,王万贤,杨毅,等.枫杨林下八种主要植物对钉螺分布的影响[J].湖北大学学报(自然科学版),1999,21(2):171~173.
[19]汪奇志,汪天平,葛继华,等.钉螺生存发育与土壤成分的相关研究[J].热带病与寄生虫学,2001,30(2):72 ~116.
[20]熊晓娇,刘立德,宋亚莉,等.滩地林分郁闭度与钉螺密度关系的研究[J].湖北林业科技,2006,(总第138 期):14 ~16.
[21]张家来,冯延寿,魏天钦,等.江滩林农复合生态系统与钉螺分布关系的研究[J].应用生态学报,1996,7(增刊):51 ~56.
[22]张旭东,漆良华,黄玲玲,等.山丘区土壤环境因子对钉螺(Oncamelania Snail)分布的影响[J].生态学报,2007,27(6):2460~2467.
[23]朱中亮.我国钉螺地理分布规律的研究[J].动物学杂志,1992,27(3):6 ~9.
[24]胡相.自然环境与钉螺壳形的变异[J].化石,1993,(1):17~19.
[25]张垒,庄黎,陈婧,等.湖北钉螺在不同温度下的发育速率及形态变化[J].湖北大学学报(自然科学版),2008,30(2):205~207.
[26]梁幼生,缪峰名,肖荣炜,等.钉螺在不同纬度地区生存繁殖的研究[J].中国血吸虫病防治杂志,1996,8(5):259 ~262.
[27]王经邦,刘寿喜.钉螺壳体“角度”与地区性差异的研究[J].科学通报,1965a,(9):812 ~814.
[28]王经邦,刘寿喜.钉螺壳体“斜高”与地区性差异的研究[J].科学通报,1965b,(9):814 ~815.
猜你喜欢
热带病与寄生虫学(2022年4期)2022-09-20
减速顶与调速技术(2022年3期)2022-02-11
热带病与寄生虫学(2022年6期)2022-02-11
落叶果树(2021年6期)2021-02-12
湖北大学学报(自然科学版)(2021年1期)2021-01-06
制造技术与机床(2019年4期)2019-04-04
广东技术师范大学学报(2016年5期)2016-08-22
体育科技(2016年2期)2016-02-28
焊接(2016年2期)2016-02-27
西华师范大学学报(自然科学版)(2015年3期)2015-02-27
