细胞自噬的研究进展
2012-11-20 01:31孙雅婧郭青龙
药学与临床研究 2012年3期
孙雅婧,郭青龙
中国药科大学生理教研室,南京 210009
细胞自噬(autophagy)是指细胞内受损、变性或衰老的蛋白质和细胞器被运输到溶酶体,溶酶体对其消化降解,以胞质内自噬体的出现为标志的细胞自我消化过程,以双层膜结构包裹部分胞质和细胞器的自噬体为判断指标。早在1962年,自噬现象的奠基人Ashford和Porten在人的肝细胞中用电子显微镜观察到了自噬现象。随着分子生物技术的发展,人们对自噬的形态特点和分子机制了解逐步深入。近年来对自噬的研究十分广泛,自噬是在体内普遍存在的过程,其在清除代谢废物进而回收能量为细胞正常运转提供能量的过程中发挥重要作用,因而对自噬的研究尤为重要。
1 细胞自噬的研究现状
1.1 自噬的过程
自噬的过程分为四个阶段(见图1)。
第一阶段:自噬诱导信号被细胞接受后,类“脂质体”碗状结构即在胞浆某处形成小的膜结构,在电镜下观察到其不断扩张、呈非球形、扁平状双层膜的碗状结构,称为自噬前体(phagophore),这种结构的电镜观察结果是指示自噬发生的金标准之一。
第二阶段:不断延伸的自噬前体,将胞浆中的若干成分(包括细胞器)收口包入,成为密闭的球状自噬体(autophagosome)。自噬体的电镜观察结果是指示自噬发生的金标准之一。自噬体的特征有两个:双层膜,内含诸如线粒体、内质网碎片等胞浆成分。
第三阶段:自噬体形成后,可能与细胞内吞的吞噬泡(phagocytic vacuole)、吞饮泡(pinosome)和内体(endosome)融合(此阶段为非必需步骤)。
第四阶段:自噬体与溶酶体(lysosome)发生融合,形成自噬溶酶体(autolysosome)。期间溶酶体酶降解自噬体的内膜,使两者的内容物合为一体,自噬体中的包含物被降解,将产物诸如氨基酸、脂肪酸之类输送到胞浆中,重新利用供能,残渣则被排出细胞外或滞留于胞浆[1]。
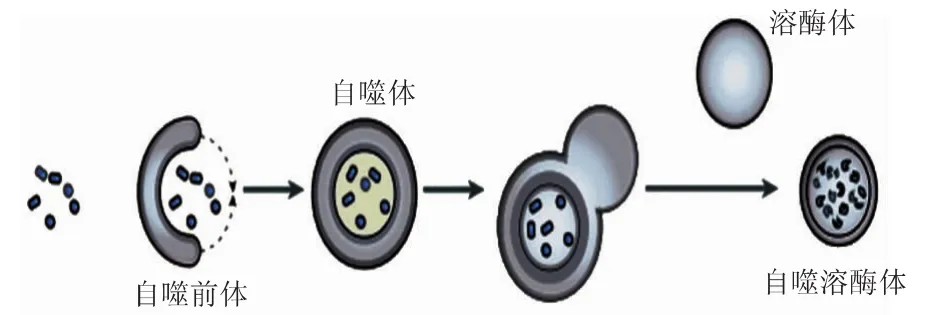
图1 自噬的基本过程
1.2 自噬的分类
根据细胞内底物运送到溶酶体腔方式的不同,哺乳动物细胞自噬分为3种主要方式:巨自噬(macroautophagy)、微自噬(microautophagy)和分子伴侣介导自噬(chaperone-mediated autophagy,简称CMA)。巨自噬是最主要的自噬形式,在巨自噬中由内质网来源的膜包绕待降解物,形成自噬体后与溶酶体融合并降解其内容物;然而在微自噬中,溶酶体膜直接内陷包裹长寿命蛋白等,并在溶酶体内降解,没有形成自噬小体的过程;分子伴侣介导自噬则为胞浆内蛋白结合到分子伴侣后转运到溶酶体腔中,被溶酶体酶消化。CMA的底物是可溶蛋白分子,因此CMA降解途径在清除蛋白质时有选择性,而前两者无明显的选择性[3]。
2 自噬与凋亡
在多细胞生物体内,维持自身的稳态和内环境的平衡,是保持复杂生物体系正常运转的重要条件。正常的细胞体系当中,有细胞的生长增殖必然伴随着细胞的衰老和死亡,细胞死亡包括3种类型:坏死、凋亡和自噬性细胞死亡。活体内局部组织、细胞的死亡称为坏死(necrosis);坏死组织细胞的代谢停止,功能丧失;坏死的形态变化可由损伤细胞内的水解酶降解作用引起,也可由游走的白细胞释放水解酶作用引起,以细胞核的改变为形态学标志。凋亡,即Ⅰ型程序性细胞死亡的特征,主要有依赖一系列蛋白水解酶即胱冬肽酶(caspase)参与,染色体浓聚、细胞皱缩、DNA降解和凋亡小体形成等,其细胞的残余部分最终被巨噬细胞清除。自噬性细胞死亡,即Ⅱ型程序性细胞死亡,以自噬体的出现为特征,不依赖于一系列蛋白水解酶即胱冬肽酶的参与,自噬体和其内的成分最终通过自身溶酶体系统被清除[4]。因此,自噬和凋亡无论在生化代谢途径,还是形态学方面都有显著区别,但研究证实,两者在功能上存在联系:在诱导细胞死亡的过程中,细胞凋亡和自噬保持着动态平衡,即自噬可能为凋亡所需,自噬通常先于凋亡,进而启动凋亡;自噬亦可能抑制凋亡作用,可保护细胞免于发生凋亡和坏死;自噬还可能向凋亡转化,共同促进细胞死亡。
大量证据还表明,自噬可能延迟凋亡的发生,抑制自噬作用可能诱发凋亡现象,同时抑制凋亡作用,也可能导致自噬现象的发生[5-6]。
3 自噬与肿瘤
自噬现象在肿瘤发生发展的过程中发挥着“双刃剑”的作用。自噬对于肿瘤的具体影响不可一概而论,不能简单将其划分为“有益的”和“有害的”,据研究表明,自噬对于肿瘤的影响是分阶段和有针对性的。
3.1 自噬对肿瘤的抑制作用
正常的多细胞生物体中,进行正常的能量代谢和生物活动同时,还会产生一些不必要的老化细胞器或蛋白及代谢物,自噬在清除代谢废物进而回收能量,为细胞正常运转提供能量的过程中发挥着重要作用。如果自噬作用受影响,细胞一些正常代谢活动不能正常进行,生物体自发性肿瘤的发生几率便可能增多。研究发现,肿瘤发生与自噬缺陷有关:如自噬相关基因BECN为杂合子的小鼠发生自发性恶性肿瘤的风险增高;乳腺癌、卵巢癌和前列腺癌以及白血病患者中,Beclin-1等位基因缺失的比例高于健康人[7-9]。在部分胶质瘤患者当中,Beclin-1蛋白 (BECN编码蛋白)和微管相关蛋白质轻链3(microtubule-associated protein 1 light chain 3,MAP-LC3)的表达均降低[10]。
其他与自噬相关基因的变化情况也有研究涉及。例如,在自然杀伤性白血病中的自噬相关基因5(autophagy-related gene5,Atg5), 在胃癌和结直肠癌中的紫外线抵抗相关肿瘤抑制基因(UV radiation resistance-associated gene,UVRAG),人胰管癌中的肿瘤抑制基因Bax作用子 (Bax-interacting factor-1,Bif-1)[11-13]。总之,基础性的自噬对于癌症发生的早期阶段起着积极的对抗作用。
3.2 自噬对肿瘤的促进作用
当肿瘤已经发生,在肿瘤细胞中出现由于严重代谢压力造成的自噬时,肿瘤细胞的数量会不断增加。自噬为生活环境恶劣的肿瘤细胞提供了有效的供养途径,使肿瘤细胞能够在低氧和低养分的微环境、尤其是缺乏血液供氧和供养分的环境下继续生存,使得肿瘤能够抵抗外界压力进行发生侵袭和转移[14]。细胞与胞外基质或者毗邻细胞脱离接触而诱发的特殊细胞程序性死亡称之为 “失巢凋亡(anoikis)”,而肿瘤细胞能够克服这种失巢凋亡,在血管缺失的情况下继续存活,而自噬作用是其作用机制之一[15]。多项体内和体外研究表明,在脱离细胞外基质、缺氧、缺养分的情况下,凋亡缺陷的肿瘤细胞表现出惊人的逆境生存能力,这种强大的生存能力与自噬机制密切相关。自噬为肿瘤细胞提供庇护,通过多种途径抑制自噬可能将提高肿瘤对放疗、化疗、免疫治疗的敏感性[16-18]。
4 自噬在抗癌药物介导的细胞死亡中发挥重要的作用
鉴于自噬在肿瘤中特殊的作用和地位,研究其在肿瘤发生发展中的分子调控至关重要。在正常细胞中,雷帕霉素靶蛋白 (mammalian target of rapamycin,mTOR) 作为磷脂酰肌醇 3-激酶(phosphatidylinositol 3-kinase,PI3K)的下游分子,在自噬中起着最初的调节作用。在生长因子的作用下,活化的PI3K/AKT/mTOR轴抑制自噬并刺激细胞的增殖。相反,在饥饿状况下PI3K通路被抑制,自噬被激活,细胞内的各种能量代谢过程受到影响,代谢处于短暂快速供能以维持细胞基本存活需要。肿瘤细胞中的自噬调节机理类似,但是过程和方式更为复杂:在肿瘤细胞中的PI3K异常激活,PI3K/AKT/mTOR途径和其他细胞信号传导级联之间的相互作用失衡,从而引起了细胞内多种异常状况的发生[19]。基于这种信号转导的异常,自噬在抗癌药物介导的细胞死亡中发挥着重要的作用,主要有以下两个方面:
4.1 自噬与凋亡的动态平衡
Hanahan和Weinberg将自噬和坏死与凋亡并称为细胞死亡的几种方式[20],但自噬和凋亡的相互关系未有定论。另有研究指出,自噬和凋亡可能受共同的上游信号调控,可能被同一个关键分子调节,Beclin-1是其中一个关键分子[21]。因而对于化疗药物所诱导的自噬和凋亡的研究相对具有针对性,但是在不同的药物作用于不同的细胞株当中,所诱导的自噬现象发挥的作用也不尽相同:抗凋亡、促生长,甚至导致细胞死亡。
化疗药物诱导自噬的机制研究未有定论,化疗药物如DNA损伤剂、微管干扰分子和激酶抑制剂也因其种类不同诱导自噬的机制也有所不同。比较公认的通路是P53的活化在转录水平上增加了调节自噬通路的相关蛋白,如腺苷单磷酸活化蛋白激酶(AMP-activated protein kinase,AMPK)、凋亡相关蛋白激酶1(death-associated protein kinase 1,DAPK1)、结节性硬化蛋白 2(tuberous sclerosis protein 2,TSC2)、unc-51 样激酶 (unc-51-like kinase 1/2,ULK 1/2)和 SESN 基因编码蛋白 1/2(sestrin 1/2)的表达[22]。
自噬与凋亡共同参与各种抗癌化疗方式诱导的细胞死亡,在某一些实例中完整的自噬机制是细胞死亡的必需步骤。在不同的细胞株,诸如肝癌细胞[23]、宫颈癌细胞[24]、甲状腺肿瘤细胞[25]等给予不同的化疗药物后,自噬可能扮演了促进细胞死亡的作用。上述研究证明,凋亡缺失的细胞中自噬性死亡成为细胞死亡的主要途径。
体内外实验的结果均证实,肿瘤组织的缺氧环境影响着抗癌药物的作用效果[26]。出现这种抵抗的原因可能是对凋亡的抑制,从而引起了缺氧诱导因子(hypoxia inducible factor-1,HIF-1)转录水平活化。随着越来越多的研究涉及自噬参与化疗药物引起细胞死亡,更多研究聚焦于抗癌药物引起的自噬中缺氧情况的变化。肿瘤组织中的持续缺氧可能扰乱凋亡和自噬两者间在细胞死亡中的平衡状态,进而引起一系列的变化[27]。
4.2 自噬的细胞保护作用
自噬作用能使肿瘤细胞在恶劣的微环境中得以生存,并使肿瘤细胞对化疗药物产生耐药作用。在某些给予抗肿瘤药的肿瘤细胞中检测到了自噬现象的大量发生[28]。在那些细胞中给予自噬抑制剂可更能有效地提高抗癌药物的治疗效果,具体机制研究尚无定论。
自噬抑制可减少肿瘤细胞对抗癌药物的抵抗机制,可能源于细胞死亡中的一个关键分子开关。濒死细胞释放损害关联模式分子(damage-associated molecular pattern molecules,DAMPs),其中一种称为高迁移率族蛋白1(high mobility group box 1 protein,HMGB1)。这种蛋白通过晚期糖化终产物受体 (receptor for advanced glycation endproducts,RAGEs),作用于其他分子诱导自噬和耐药。此作用途径目前仅发现于白血病细胞[29],其他肿瘤细胞尚未有相关研究。
5 自噬现象的临床意义
随着自噬现象研究的深入,自噬渐渐成为一个新的癌症治疗靶点。如前所述,半数研究表明,自噬是抗癌治疗中有效杀伤肿瘤细胞所必需的。研究者期望设计出新的药物诱导肿瘤细胞的自我吞噬,甚至寄希望于自噬可以清除肿瘤细胞。某些以通过诱导自噬达到影响肿瘤细胞作用的药物将要作进一步的临床试验[30]。
化疗药物使用后,肿瘤细胞中自噬现象的大量产生预示着一种细胞的自我保护反应,以自噬体的形成与融合为靶点的治疗药物为肿瘤化疗提供了一条新思路和新途径[31]。因而诱导肿瘤细胞发生自噬的药物,在肿瘤的预防和治疗初期阶段有着广阔的前景和极其重要的意义。
[1]Mizushima N.Autophagy:process and function[J].Genes Dev,2007,21(22):2861-73.
[2]Levine B.Cell biology:autophagy and cancer[J].Nature,2007,446(7137):745-7.
[3]Yoshimori T.Autophagy:a regulated bulk degradation process inside cells[J].Biochem Biophys Res Commun,2004,313(2):453-8.
[4]Lockshin RA,ZakeriZ.Apoptosis,autophagy,and more[J].Int J Biochem Cell Biol,2004,36(12):2405-19.
[5]Maiuri MC,Zalckvar E,Kimchi A,et al.Self-eating and self-killing:crosstalk between autophagy and apoptosis[J].Nat Rev Mol Cell Biol,2007,8(9):741-52.
[6]Platini F,Perez-Tomas R,Ambrosio S,et al.Understanding autophagy in cell death control[J].Curr Pharm Des,2010,16(1):101-13.
[7]Qu X,Yu J,Bhagat G,et al.Promotion of tumorigenesis by heterozygous disruption of the beclin 1 autophagy gene[J].J Clin Invest,2003,112(12):1809-20.
[8]Yue Z,Jin S,Yang C,et al.Beclin 1,an autophagy gene essential for early embryonic development,is a haploinsufficient tumor suppressor[J].Proc Natl Acad Sci USA,2003,100(25):15077-82.
[9]Liang XH,Jackson S,Seaman M,et al.Induction of autophagy and inhibition of tumorigenesis by beclin 1[J].Nature,1999,402(6762):672-6.
[10]Huang X,Bai HM,Chen L,et al.Reduced expression of LC3B-II and Beclin 1 in glioblastoma multiforme indicates a down-regulated autophagic capacity that relates to the progression of astrocytic tumors[J].J Clin Neurosci,2010,17(12):1515-9.
[11]Iqbal J,Kucuk C,Deleeuw RJ,et al.Genomic analyses revealglobalfunctionalalterationsthatpromote tumor growth and novel tumor suppressor genes in natural killer-cell malignancies[J].Leukemia,2009,23(6):1139-51.
[12]Liang C,Feng P,Ku B,et al.Autophagic and tumour suppressor activity of a novel Beclin1-binding protein UVRAG[J].Nat Cell Biol,2006,8(7):688-99.
[13]Coppola D,Khalil F,Eschrich SA,et al.Down-regulation of Bax-interacting factor-1 in colorectal adenocarcinoma[J].Cancer,2008,113(10):2665-70.
[14]Kenific CM,Thorburn A,Debnath J.Autophagy and metastasis:another double-edged sword[J].Curr Opin Cell Biol,2010,22(2):241-5.
[15]Horbinski C,Mojesky C,Kyprianou N.Live free or die:tales of homeless (cells)in cancer[J].Am J Pathol,2010,177(3):1044-52.
[16]Degenhardt K,Mathew R,Beaudoin B,et al.Autophagy promotestumorcellsurvivaland restricts necrosis,inflammation,and tumorigenesis[J].Cancer Cell,2006,10(1):51-64.
[17]Kuma A,Hatano M,Matsui M,et al.The role of autophagy during the early neonatal starvation period[J].Nature,2004,432(7020):1032-6.
[18]Pavlides S,Tsirigos A,Migneco G,et al.The autophagic tumor stroma model of cancer:role of oxidative stress and ketone production in fueling tumor cell metabolism[J].Cell Cycle,2010,9(17):3485-505.
[19]Chen N,Karantza-Wadsworth V.Role and regulation of autophagy in cancer[J].Biochim Biophys Acta,2009,1793(9):1516-23.
[20]Hanahan D,Weinberg RA.Hallmarks of cancer:the next generation[J].Cell,2011,144(5):646-74.
[21]Kang R,Zeh HJ,Lotze MT,et al.The Beclin 1 network regulates autophagy and apoptosis[J].Cell Death Differ,2011,18(4):571-80.
[22]Feng Z.p53 Regulation of the IGF-1/AKT/mTOR pathways and the endosomal compartment[J].Cold Spring Harb Perspect Biol,2010,2(2):a001057.
[23]Cosse JP,Rommelaere G,Ninane N,et al.BNIP3 protects HepG2 cells against etoposide-induced cell death underhypoxia by an autophagy-independent pathway[J].Biochem Pharmacol,2010,80(8):1160-9.
[24]Lee SB,Tong SY,Kim JJ,et al.Caspase-independent autophagic cytotoxicity in etoposide-treated CaSki cervical carcinoma cells[J].DNA Cell Biol,2007,26(10):713-20.
[25]Lin CI,Whang EE,Donner DB,et al.Autophagy induction with RAD001 enhances chemosensitivity and radiosensitivity through Met inhibition in papillary thyroid cancer[J].Mol Cancer Res,2010,8(9):1217-26.
[26]Rohwer N,Cramer T.Hypoxia-mediated drug resistance:novel insights on the functional interaction of HIFs and cell death pathways[J].Drug Resist Updat,2011,14(3):191-201.
[27]Piret JP,Mottet D,Raes M,et al.Is HIF-1alpha a pro-or an anti-apoptotic protein?[J].Biochem Pharmacol,2002,64(5-6):889-92.
[28]Kondo Y,Kanzawa T,Sawaya R,et al.The role of autophagy in cancerdevelopmentand response to therapy[J].Nat Rev Cancer,2005,5(9):726-34.
[29]Liu L,Yang M,Kang R,et al.DAMP-mediated autophagy contributes to drug resistance[J].Autophagy,2011,7(1):112-4.
[30]Levy JM,Thorburn A.Targetingautophagy during cancer therapy to improve clinical outcomes[J].Pharmacol Ther,2011,131(1):130-41.
[31]Chen S,Rehman SK,Zhang W,et al.Autophagy is a therapeutic targetin anticancerdrug resistance[J].Biochim Biophys Acta,2010,1806(2):220-9.
猜你喜欢
中国交通信息化(2022年8期)2022-10-28
中国种业(2022年9期)2022-10-13
九江学院学报(自然科学版)(2022年2期)2022-07-02
中国兽医学报(2022年4期)2022-06-17
河南科技(2022年6期)2022-04-22
小学生学习指导(低年级)(2021年6期)2021-07-19
小学生学习指导(中年级)(2021年6期)2021-06-19
小学生学习指导(高年级)(2021年6期)2021-06-19
现代临床医学(2021年1期)2021-01-26
好日子(2018年9期)2018-10-12