
人工老化诱导的油菜种子活力和抗氧化酶活性变化的研究
2012-11-12 00:59:10曾钦薇谢永俊
云南农业科技 2012年2期
曾钦薇,谢永俊
(1.云南省农业科学院蚕桑蜜蜂研究所,云南蒙自661101;2.云南省农业科学院经济作物研究所,云南昆明650205)
植物种子长期贮存期间活力将逐渐丧失,但老化的速率因植物种类而异[1]。老化原因及其抗衰老机制国内外已作了一些研究。老化期间,种子产生大量活性氧中间体(reactive oxygen species,ROS),ROS导致脂质过氧化[2],丙二醛(MDA)则是脂质过氧化的产物,其含量的高低在一定程度上反映了膜脂过氧化的水平[3]。大量研究表明,自由基是种子老化的主要原因,它导致脂质过氧化,对细胞膜结构的完整性和功能造成巨大伤害,使种子活力衰退。种子拥有包括自由基和过氧化物清除酶在内的一些保护机制,这些酶如过氧化氢酶(CAT)、过氧化物酶(POD)、抗坏血酸过氧化物酶(APX)、谷胱甘肽还原酶(GR)和超氧化物歧化酶(SOD)[2,4]。以向日葵为例,种子的老化与去氧化酶活性的降低和脂质过氧化密切相关[5]。研究还表明,种子高含水量和高温将进一步加速种子衰老[6]。
甘蓝型油菜(Brassica napus L.)是中国重要的油料作物,在开花前期和成熟期遭受高温胁迫将影响种子产量和质量[7]。鉴于油菜种子含有较多的脂肪,贮存期间脂质可能更易被过氧化,进而加速种子活力衰退,本研究通过高温高湿处理加速新鲜油菜种子老化,测定其活力、抗氧化酶活性和MDA含量等的变化,进一步探讨种子活力衰退的生理生化机制。
1 材料与方法
1.1 主要仪器设备
电热恒温水浴锅(HHS型,上海博讯),电导仪(JEY-45001,Jenway,UK),梅特勒XS105DU电子天平(瑞士),高速冷冻离心机(Eppendorf centrifuge 5810R,Germany),双光束紫外可见分光光度计(TU-1901,北京普析)。
1.2 油菜种子
产自云南省弥勒县的甘蓝型油菜新鲜种子,品种为花油7号。
1.3 种子老化处理
参考Parrish and Leopid方法[8],稍作改进。每个处理选取100粒饱满的种子,加5%次氯酸钠溶液涡旋消毒3 min,去离子水漂洗3次,用滤纸吸干;转入平皿,摊平,置45℃水浴,水浴锅加盖密封(使相对湿度保持100%)。老化处理1 d、2 d、3 d,测定各生理指标。
1.4 测定方法
1.4.1 种子发芽指数、活力指数、发芽率和发芽速率测定
采用GB/T3543.4-1995技术规范测定种子发芽率、发芽指数、活力指数和发芽速率[9]。老化时间分别为0 d(CK)、2 d、4 d、6 d,3个重复,每个处理50粒种子。发芽指数(GI)=∑(Gt/Dt),活力指数(VI)=GI×S,发芽率(%)=发芽终期全部正常种苗数/供试种子数×100,发芽速率(日)=∑(Gt×Dt)/发芽率。式中,Gt为每天的发芽粒数,Dt为相应发芽日数,S为观察结束时幼苗长度(根长+茎长,cm)。
1.4.2 种子相对电导率的测定
种子发生劣变时,细胞膜结构和功能完整性受到伤害,质膜透性增加。透性增加将使种子向周围介质溶液渗漏出较多的电解质,使溶液电导率增加,电导率的高低则反映出种子质膜的受损程度,受损越严重,种子活力越低。仿Zhang等方法[10],并适当改进。取经不同时间[0 d(CK)、1 d、2 d、3 d]加速老化处理的5粒种子放入盛有5 ml去离子水的具塞试管内,置于25℃/120 rpm摇床振荡1 h,测定初电导率(ECi);试管转入100℃沸水浴中加热10 min,流水冷却到室温,测定终电导率(ECt)。以ECi/ECt表示相对电导率。3个重复。
1.4.3 MDA测定
参照Dhindsa等硫代巴比妥酸法[11]。3个重复。
1.4.4 POD、SOD和CAT活性测定
采用陈建勋等方法[12]测定CAT、POD和SOD活性。3个重复。
1.5 统计分析
采用SPSS 17.0统计软件进行统计分析。
2 结果与分析
2.1 老化处理对油菜种子发芽指数、活力指数、发芽率和发芽速率的影响
从表1可见,油菜种子的发芽指数、活力指数、发芽率和发芽速率随着老化时间的延长而降低,前3个指数各老化时间之间及其与对照组之间差异极显著(p<0.01)。发芽速率与对照组差异也极显著(p<0.01)。显然,加速老化使油菜种子的活力大大降低。
表1 老化处理油菜种子的发芽指数、活力指数、发芽率和发芽速率(n=3)

表1 老化处理油菜种子的发芽指数、活力指数、发芽率和发芽速率(n=3)
项目 老化时间(d)0(CK)1 2 3发芽指数 41.5±8.6A 20.4±5.7B 13.2±3.4C 1.1±0.3D活力指数 256.9±33.4A 67.7±13.6B 28.8±9.3C 2.8±1.1D发芽率(%)99.7±0.1A 88.1±3.3B 64.8±5.3C 8.2±2.2D发芽速率(日)0.7±0.1A 1.3±0.1B 1.5±0.2C 2.1±0.2D
表2 老化处理对油菜种子的相对电导率和生化指标的影响(,n=3)
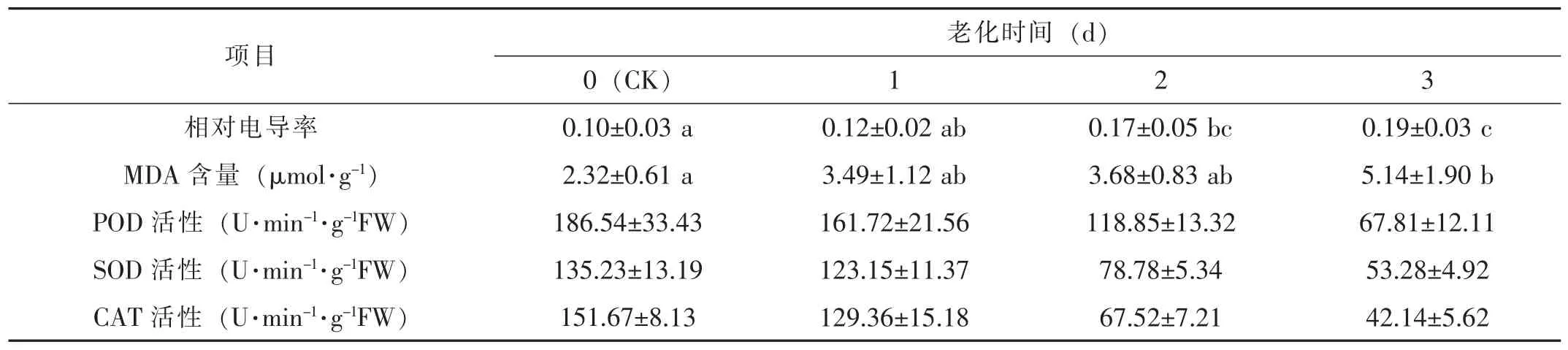
表2 老化处理对油菜种子的相对电导率和生化指标的影响(,n=3)
老化时间(d)0(CK)1 2 3相对电导率 0.10±0.03 a 0.12±0.02 ab 0.17±0.05 bc 0.19±0.03 c项目MDA 含量(μmol·g-1)2.32±0.61 a 3.49±1.12 ab 3.68±0.83 ab 5.14±1.90 b POD 活性(U·min-1·g-1FW)186.54±33.43 161.72±21.56 118.85±13.32 67.81±12.11 SOD 活性(U·min-1·g-1FW)135.23±13.19 123.15±11.37 78.78±5.34 53.28±4.92 CAT 活性(U·min-1·g-1FW)151.67±8.13 129.36±15.18 67.52±7.21 42.14±5.62
2.2 老化处理对油菜种子细胞膜完整性的影响
从表2可见,种子渗出液的相对电导率随着老化时间的延长而增加。方差分析表明,老化2 d、3 d与对照、老化1 d与3 d差异显著(p<0.05),但老化1 d与对照、老化2 d与老化3 d差异不显著(p>0.05)。显然,老化时间越长,油菜种子细胞膜的完整性被破坏越大,将降低种子的活力。
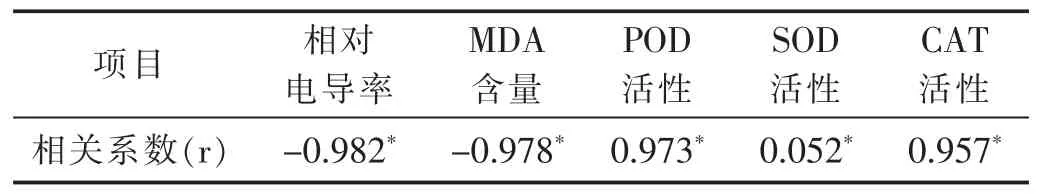
表3 老化处理油菜种子活力与各生理指标之间的相关性
2.3 老化处理对油菜种子MDA含量的影响
随着老化时间的延长,MDA含量呈逐渐上升趋势(表2)。方差分析表明,老化3 d与对照组差异显著(p<0.05),其余各组之间差异不显著(p>0.05)。显然,老化时间越长,油菜种子中的脂质更多地被过氧化,进而加速种子活力衰退。
2.4 老化处理对油菜种子POD、SOD和CAT活性的影响
随着老化时间的延长,油菜种子中POD、SOD和CAT的活性呈下降趋势(表2),老化时间越长,与对照的差异越大。显然,随着老化时间的延长,种子内上述过氧化物清除酶的活性降低,脂质将更多地被过氧化,使种子活力下降。
2.5 种子活力与上述各生理指标之间的相关性分析
采用SPSS17.0统计软件进行相关性分析表明,油菜种子活力与相对电导率和MDA含量呈显著负相关,而与POD、SOD和CAT活性(表3)呈显著正相关(p<0.05)。
3 讨论
种子中自由基的产生将加速脂质过氧化,并破坏细胞膜结构和功能的完整性。所产生的自由基如果不能被清除,必将因脂质过氧化、细胞膜结构和功能完整性的破坏,使种子老化,活力降低。细胞中产生的超氧化物自由基和膜脂中的不饱和脂肪酸发生过氧化是膜被破坏的原因[3]。因此,种子中的自由基和过氧化物清除酶—SOD、POD、CAT、APX、GR在种子抗衰老中的作用十分重要。SOD催化2分子过氧化物为分子氧和H2O2[13]。CAT、POD、APX与 H2O2清除有关,APX用抗坏血酸作为电子供体将H2O2还原为水[14],GR通过谷胱甘肽—抗坏血酸氧化还原循环在内生H2O2控制中起作用[15],而且,抗坏血酸和谷胱甘肽在植物中是重要的抗氧化剂[16]。
在本研究中,加速老化使油菜种子中POD、SOD和CAT活性及质膜的完整性降低,过氧化产物MDA含量增加,外在表现为种子发芽指数、活力指数、发芽率和发芽速率下降。本研究与Afshari等[17]测定的人工老化油菜种子结果相似。POD活性的变化与王煜等[18]测定的自然老化19年的油菜种子的结果相一致。
综上所述,油菜种子老化期间各种抗氧化酶活性的下降、脂质的过氧化、细胞膜结构和功能完整性的破坏,是其老化和活力衰退的原因。鉴于高温高湿是诱导种子老化的重要外因,在贮存油菜种子时应将其含水量降至安全范围,同时要降低贮存温度,以延长种子寿命。
[1]Dhakal M R,Pandey A K.Storage potential of niger(Guizotia abyssinica Cass.)seeds under ambient conditions[J].Seed Sci.Technol.,2001,29:205~213.
[2]McDonald M B.Seed deterioration:physiology,repair and assessment[J].Seed Sci.Technol.,1999,27:177~237.
[3]娜荷雅.高温对油菜、燕麦和大豆种子生理代谢及衰老的影响[A].呼和浩特:内蒙古农业大学,2008.
[4]Bowler C,van Montagu M,Inze D.Superoxide dismutase and stress tolerance[J].Annu.Rev.Plant Physiol.Plant Mol.Biol.,1992,43:83~116.
[5]Bailly C,Benamar A,Corbineau F,et al.Changes in malondialdehyde content and in superoxide dismutase,catalase and glutathione reductase activities in sunflower seeds as related to deterioration during accelerated ageing[J].Physiologia Plantarum,1996,97:104~110.
[6]Abba E J,Lovato A.Effect of seed storage temperature and relative humidity on maize(Zea mays L.)seed viability and vigour[J].Seed Sci.Technol.,1999,27:101~114.
[7]汪剑鸣,龚乃弘.气象因子与油菜产量关系的初步研究[J].江西农业学报,1997,9(1):6~11.
[8]Parrish D J,Leopold A C.On the mechanism of aging in Soybean seeds[J].Plant Physiol.,1978,61:365~368.
[9]毕辛华,戴心维.种子学[M].北京:中国农业出版社,2002.
[10]Zhang J,Huang W,Pan Q,et al.Improvement of chilling tolerance and accumulation of heat shock proteins in grape berries(Vitis vinifera cv.Jingxiu)by heat pretreatment[J].Postharvest Biology and Technology,2005,38:80~90.
[11]Dhindsa R S,Plumb-Dhindsa P,Thorpe T A.Leaf senescence:correlated with increased leaves of membrane perme-ability and lipid peroxidation and decreased levels of superoxide dismutase and catalase[J].J.Exp.Bot.,1981,32:93~101.
[12]陈建勋,王晓峰.植物生理学实验指导(第二版)[M].广州:华南理工大学出版社,2006.
[13]Scandalios J G.Oxygen stress and superoxide dismutase[J].Plant Physiol.,1993,101:7~12.
[14]Nakano Y,Asada K.Hydrogen peroxide is scavenged by ascorbate-specific peroxidase in spinach chloroplasts[J].Plant Cell Physiol.,1981,22:867~880.
[15]Smith I K,Vierheller T L,Thorne C.Properties and functions of glutathione reductase in plants[J].Physiol.Plant,1989,77:449~456.
[16]Tommasi F,Paciolla C,Arrigoni O.The ascorbate system in recalcitrant and orthodox seeds[J].Physiol.Plant,1999,105:193~198.
[17]Afshari R T,Rashidi S,Alizadeh H.Effects of seed aging on germination characteristics and on catalase and peroxidase activities in two canola cultivars(Brassica napus L.)[J].Iranian Journal of Field Crop Science,2009,40(2):125~133.
[18]LI Li,HUANG Xian-qun,Wanapu Chokchai,et al.Assessment of Genetic Diversity and Relationships among Brassica napus Using AFLP Markers[J].西南农业学报,2010,23(4):1243~1251.
[19]王煜,田廷亮,扶惠作,等.油菜种子老化过程中的生理生化变异[J].中国油料,1994,16(3):11~14.
猜你喜欢
今日农业(2022年6期)2022-07-05 15:13:21
现代临床医学(2021年6期)2021-11-20 06:34:50
科学导报(2020年8期)2020-03-12 04:51:04
中国食品学报(2020年2期)2020-01-17 07:59:08
种子科技(2018年11期)2018-09-10 00:56:48
许昌学院学报(2018年8期)2018-09-05 02:05:18
中成药(2017年12期)2018-01-19 02:06:48
潍坊学院学报(2016年2期)2016-12-01 12:59:44
天津农林科技(2015年1期)2015-12-30 13:05:14
中国洗涤用品工业(2015年4期)2015-02-28 19:02:15
