
跑台运动对比目鱼肌和腓肠肌肌肉生长抑制素表达的影响
2012-11-09 05:42刘小园杨洪涛
沈阳体育学院学报 2012年5期
张 靓,刘小园,杨洪涛
(北京师范大学体育与运动学院,北京 100875)
◂运动人体科学
跑台运动对比目鱼肌和腓肠肌肌肉生长抑制素表达的影响
张 靓,刘小园,杨洪涛
(北京师范大学体育与运动学院,北京 100875)
目的:分别观察运动促进的骨骼肌肥大时,比目鱼肌和腓肠肌肌肉生长抑制素(myostatin,MSTN)表达的变化。方法:16周龄雄性SD大鼠随机分为对照组和运动组,每组10只。跑台训练持续8周,每周5次,每次60min,跑速26m/min。测定两组大鼠肌重胫骨长的比值,HE染色观察骨骼肌纤维横截面积,分别采用real-time PCR技术和western blot方法测定比目鱼肌和腓肠肌MSTN mRNA和蛋白表达。结果:运动大鼠骨骼肌重量/胫骨长显著升高(P<0.05),两种肌纤维横截面积均增大。与对照组相比,跑台运动显著抑制了大鼠腓肠肌MSTN的mRNA和蛋白表达(P<0.05),但对比目鱼肌MSTN表达的影响无显著性。结论:运动对MSTN的作用可能存在肌纤维特异性。
肌肉生长抑制素;肌肉肥大;比目鱼肌;腓肠肌
肌肉生长抑制素(myostatin,MSTN),即生长分化因子8(growth differentiation factor 8,GDF-8),为转化生长因子 β(transforming growth factor beta,TGFβ)超家族成员之一,具有与TGFβ超家族的典型结构特点。MSTN基因先合成含375个氨基酸的前体蛋白,经二次蛋白酶酶解活化,首先切除23个氨基酸的信号肽形成前肽,之后在第263-266位被裂解,形成N端约243个氨基酸和C端109个氨基酸的多肽,后者以二硫键的方式结合成二聚体,与膜上的特异性受体结合,发挥生物学作用[1]。MSTN在骨骼肌大量表达,并因其对骨骼肌生长具有强大的负调控能力而备受关注[2]。大量研究证实,不管是在 MSTN表达缺失的动物[3-4],还是在 MSTN突变的人群[5],均出现了异常的肌肉肥大。此外,后天使用MSTN抑制剂[6]、MSTN抗体或使用短发卡RNA干扰MSTN的表达后[7],也同样诱导了肌肉质量的增加。与此相反,当过表达MSTN时则诱导小鼠骨骼肌萎缩[8-9],并且在肌肉萎缩的动物模型[10]和临床肌肉萎缩的患者[11-12]均出现骨骼肌中MSTN表达的显著增强,提示MSTN参与了骨骼肌结构和功能的生理和病理生理调节。
但目前,越来越多的研究提示,MSTN对骨骼肌的作用存在肌纤维特异性。临床研究发现,在多种以II型肌纤维萎缩为主的肌肉萎缩性疾病,如恶变质时的肌肉萎缩、长期糖皮质激素治疗引起的肌肉萎缩等,均发现骨骼肌MSTN表达显著升高[13]。在食物剥夺诱导的小鼠II型纤维肌肉萎缩模型上[14],发现胫骨前肌出现了MSTN mRNA表达的显著上调;在去神经诱导的肌萎缩大鼠模型[15],同样是在腓肠肌中MSTN表达上调,并且与腓肠肌的萎缩程度呈显著相关。而在大鼠悬挂模型中,比目鱼肌湿重减轻42%,腓肠肌湿重减轻17%,但只有在腓肠肌中MSTN mRNA水平升高67%,提示MSTN的表达不与骨骼肌的萎缩程度相关,而是与萎缩的IIb型肌球蛋白重链显著相关[16]。此外,在MSTN基因敲除的小鼠,骨骼肌肌纤维类型也向糖酵解型肌纤维方向转变[17-19]。上述研究提示,MSTN的表达变化对 II型肌纤维有着特异性的调节作用。
大量文献报道了运动对骨骼肌MSTN表达的影响,但长期的有氧运动对不同肌纤维中MSTN的mRNA和蛋白表达水平的影响尚不明确。本研究观察长期的跑台运动对大鼠比目鱼肌(I型肌纤维为主)和腓肠肌(II型肌纤维为主)中MSTN的mRNA和蛋白表达的变化,探究运动对MSTN的作用是否也存在肌纤维特异性。
1 材料与方法
1.1 实验动物与材料
实验所有动物饲养和实验程序按中国动物管理条例(中华人民共和国卫生部2001年第55号文件)批准的动物实验标准进行。16周龄雄性SD大鼠由北京大学医学部动物中心提供,实验动物生产许可证号:SCXY(京)2002-000。常规条件下饲养,室内温度为(22±2)℃,湿度为40% ~60%,自由进食和饮水。MSTN抗体及β-actin抗体均自Santa Cruz Biotechnology(Santa Cruz,CA,USA)。实验中所用其他试剂系市售分析纯试剂。
1.2 动物分组与运动模型建立
大鼠随机分为对照组和运动组,每组10只。运动组采用跑台运动,在第1周内,跑台速度逐渐加快至26m/min[20],跑台时间逐渐延长至60min。正式跑台训练持续8周,每周5次,每次60min。每次训练在下午3点至6点间进行,不使用光、电等刺激手段。两组动物均常规饲养8周后进行下述检测。
1.3 取材及组织准备
大鼠末次运动结束后,禁食过夜,自由饮水,腹腔注射乌拉坦(1g/kg)。放血处死动物后,分离比目鱼肌和腓肠肌,滤纸吸干称重,测量胫骨长度,用于计算肌肉重量与胫骨长度比。取少许比目鱼肌和腓肠肌(白肌部分)组织10%甲醛固定,石蜡包埋切片,HE染色,光镜下进行形态学检查。其余骨骼肌组织用锡箔纸包裹、标记后先于液氮中暂存,后转于-80℃保存,分别进行下述测定。
1.4 Real-time PCR测定骨骼肌MSTN mRNA水平
骨骼肌总RNA的提取采用Trizol一步法进行,采用Promega逆转录系统(Promega,Madison,WI,USA)进行逆转录。real-time PCR反应体积10μL:10×缓冲液1mL,10mmol/L dNTP混合物0.5mL,10mmol/L的上下游引物各0.25ml,cDNA 模板 2.5ml,Taq DNA 多聚酶 0.2ml,SYBR green I 0.4ml。MSTN的上游引物:AGC CTG AAT CCA ACT TAG;下游引物为:GTT CAT CAC AGT CAA GCC;β-actin的上游引物:5’-GAG ACC TTC AAC ACC CCA GCC -3’;下游引物为:5’-TCG GGG CAT CGG AAC CGC TCA-3’。经95℃ 7min变性后,MSTN 和 β-actin分别进行:95℃30 s,54.2℃ 30 s,72℃40 s,热循环 42 次和 95℃ 30s,54℃ 30s,72℃ 40s,热循环45次。Real-time PCR于Mx3000P多通道实时定量PCR仪(Stratagene,La Jolla,CA,USA)上进行实验。使用 β -actin作为内参。
1.5 Western-blot测定骨骼肌MSTN的蛋白表达
分别取比目鱼肌和腓肠肌(白肌部分)组织50mg,蛋白裂解液匀浆,离心后取上清,采用考马斯亮蓝法进行蛋白定量。同等蛋白量的样品经十二烷基磺酸钠-2-聚丙烯酰胺凝胶(SDS-PAGE)电泳后,转膜、5%脱脂奶粉封闭1h,MSTN(1:1000)、β -actin(1∶2000)一抗孵育,4℃过夜,二抗孵育1h,经显色后曝光。蛋白表达水平用NIH图像分析软件(Bethesda,Maryland,USA)进行分析,β-actin作为内参。
1.6 数理统计法
2 结果
2.1 跑台运动诱导大鼠比目鱼肌和腓肠肌肥大的发生
与对照组相比,8周跑台运动后,大鼠的比目鱼肌重/胫骨长和腓肠肌重/胫骨长均显著升高(P<0.05)(图1A,1B)。HE染色发现,与对照组相比,运动组大鼠比目鱼肌和腓肠肌纤维横截面积显著增大(图1C,1D),提示骨骼肌肥大的发生。
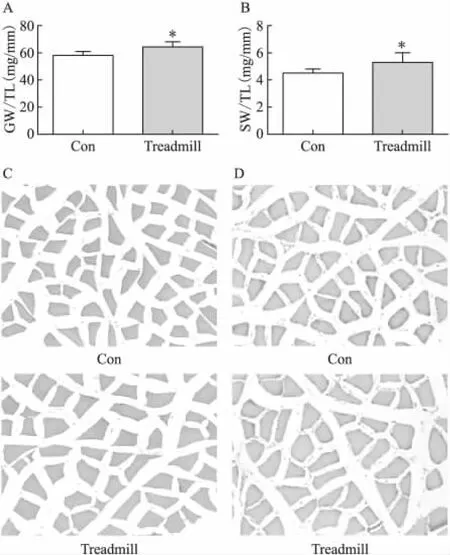
图1 跑台运动对大鼠腓肠肌/胫骨长(A),比目鱼肌/胫骨长(B),腓肠肌纤维横截面积(C)及比目鱼肌横截面积(D)的影响
2.2 跑台运动抑制大鼠腓肠肌MSTN的mRNA和蛋白表达
与对照组相比,运动组大鼠腓肠肌MSTN的mRNA表达水平和蛋白表达水平均显著降低(P<0.05)(图2A,2B)。
2.3 跑台运动不影响大鼠比目鱼肌MSTN的mRNA和蛋白表达
与对照组相比,8周的跑台运动不影响大鼠比目鱼肌MSTN的mRNA表达水平和蛋白表达水平(P>0.05)(图3A,3B)。
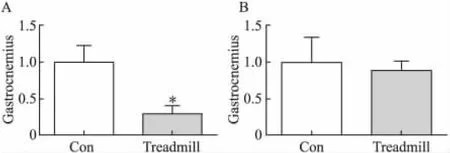
图2 跑台运动对大鼠腓肠肌和比目鱼肌MSTN mRNA表达的影响
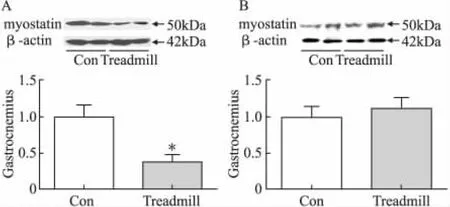
图3 跑台运动对大鼠腓肠肌和比目鱼肌MSTN蛋白表达的影响
3 讨论
肌球蛋白是构成骨骼肌纤维粗肌丝的主要成分,是骨骼肌纤维中表达最多的蛋白。天然的肌球蛋白由6条多肽链组成,包括两条重链和两对不同的轻链。肌球蛋白重链(myosin heavy chain,MHC)在成年骨骼肌有 I-MHC、Ⅱa-MHC、Ⅱx-MHC和Ⅱb-MHC 4种类型,在不同肌肉、肌肉不同区域及不同动物,MHC表达的类型和数量会有所差异,慢肌主要表达I型及部分Ⅱa型MHC,如比目鱼肌和股中间肌;快肌如腓肠肌、足底肌、股四头肌、趾长伸肌、胫骨前肌等主要以Ⅱx、Ⅱb MHC组成。在本实验中,选取I型MHC为主的比目鱼肌和IIb型MHC为主的腓肠肌,发现8周跑台运动促进了比目鱼肌和腓肠肌肥大的发生,该结果与以往的文献报道相一致[21]。
运动可通过多种机制促进骨骼肌肥大,近来骨骼肌内分泌功能的发现为运动促进骨骼肌肥大机制研究提供了新的方向。运动可通过肌肉收缩活动,调节骨骼肌旁/自分泌及内分泌功能稳态,参与骨骼肌的生长、代谢和运动功能[22]。MSTN是骨骼肌表达、合成和分泌的肌肉因子,具有强大抑制肌肉生长的作用。MSTN对骨骼肌的作用有明显的肌纤维特异性。Carlson CJ等[16]研究发现,MSTN的mRNA表达与骨骼肌中IIb MHC的含量呈线性正相关。在MSTN基因敲除的小鼠,骨骼肌中IIb MHC含量显著升高,IIa MHC的含量显著降低,与此结构变化相对应,骨骼肌中线粒体的数目、琥珀酸脱氢酶的活性均降低,疲劳易感度增加,骨骼肌的代谢向糖酵解方向转化[17-19],可见,MSTN的表达变化主要参与了IIb MHC结构和功能的调节。在实验中,发现运动显著抑制了腓肠肌中MSTN mRNA和蛋白表达,但并不影响比目鱼肌中MSTN mRNA和蛋白表达,提示在运动促进的肌肉肥大中,MSTN的作用同样也具有肌纤维类型的特异性,只参与了运动诱导的IIb型MHC肥大。
运动通过何种机制调节MSTN的表达,目前尚未见报道。Spiller MP等[23]报道,骨骼肌成肌调节因子Myo D可促进MSTN的启动子激活,上调MSTN的表达,抑制肌肉肥大。运动是否通过Myo D-MSTN途径对MSTN的表达进行转录水平调节还需更多的研究来证实。
综上所述,运动通过抑制腓肠肌MSTN的mRNA和蛋白表达,促进腓肠肌肥大的形成;运动不影响比目鱼肌MSTN的表达,运动促进的比目鱼肌肥大可能存在其他的调节机制。结果提示,在运动促进的肌肉肥大中,MSTN的作用具有肌纤维特异性。
[1]Jespersen JG,Nedergaard A,Andersen LL,et al.Myostatin expression during human muscle hypertrophy and subsequent atrophy:increased myostatin with detraining[J].Scand J Med Sci Sports,2011,21(2):215 -223.
[2]McPherron AC,Lawler AM,Lee SJ.Regulation of skeletal muscle mass in mice by a new TGF-beta superfamily member[J].Nature,1997,387:83 -90.
[3]McPherron AC,Lee SJ.Double muscling in cattle due to mutations in the myostatin gene[J].Proc Natl Acad Sci U S A,1997,94(23):12457-12461.
[4]Lee SJ.Regulation of muscle mass by myostatin[J].Annu Rev Cell Dev Biol,2004,20:61 -86.
[5]Schuelke M,Wagner KR,Stolz LE,et al.Myostatin mutation associated with gross muscle hypertrophy in a child[J].N Engl J Med,2004,350(26):2682 -2688.
[6]Morine KJ,Bish LT,Pendrak K,et al.Systemic myostatin inhibition via liver-targeted gene transfer in normal and dystrophic mice[J].PLoS One,2010(2):e9176.
[7]Magee TR,Artaza JN,Ferrini MG,et al.Myostatin short interfering hairpin RNA gene transfer increases skeletal muscle mass[J].J Gene Med,2006(9):1171 -1781.
[8]Zimmers TA,Davies MV,Koniaris LG,et al.Induction of cachexia in mice by systemically administered myostatin[J].Science,2002,296:1486-1488.
[9]Amirouche A,Durieux AC,Banzet S,et al.Down-regulation of Akt/mammalian target of rapamycin signaling pathway in response to myostatin overexpression in skeletal muscle[J].Endocrinology,2009,150(1):286-294.
[10]McFarlane C,Sharma M,Kambadur R.Myostatin is a procachectic growth factor during postnatal myogenesis[J].Curr Opin Clin Nutr Metab Care,2008,11(4):422 -427.
[11]Gonzalez-Cadavid NF,Taylor WE,et al.Organization of the human myostatin gene and expression in healthy men and HIV-infected men with muscle wasting[J].Proc Natl Acad Sci U S A,1998,95(25):14938-14943.
[12] Reardon KA,Davis J,Kapsa RM,et al.Myostatin,insulin-like growth factor-1,and leukemia inhibitory factor mRNAs are upregulated in chronic human disuse muscle atrophy[J].Muscle Nerve,2001,24(7):893 -899.
[13]Wójcik S,Nogalska A,Engel WK,et al.Myostatin and its precursor protein are increased in the skeletal muscle of patients with Type-II muscle fibre atrophy[J].Folia Morphol(Warsz),2008,67(1):6 -12
[14]Allen DL,Cleary AS,Lindsay SF,et al.Myostatin expression is increased by food deprivation in a muscle-specific manner and contributes to muscle atrophy during prolonged food deprivation in mice[J].J Appl Physiol,2010,109(3):692 -701.
[15]Zhang D,Liu M,Ding F,et al.Expression of myostatin RNA transcript and protein in gastrocnemius muscle of rats after sciatic nerve resection[J].J Muscle Res Cell Motil,2006,27(1):37 - 44.
[16]Carlson CJ,Booth FW,Gordon SE.Skeletal muscle myostatin mRNA expression is fiber-type specific and increases during hindlimb unloading[J].Am J Physiol,1999,277(2 Pt 2):R601 - 606.
[17]Girgenrath S,Song K,Whittemore LA.Loss of myostatin expression alters fiber-type distribution and expression of myosin heavy chain isoforms in slow-and fast-type skeletal muscle[J].Muscle Nerve,2005,31(1):34 -40.
[18]Amthor H,Macharia R,Navarrete R,et al.Lack of myostatin results in excessive muscle growth but impaired force generation[J].Proc Natl Acad Sci U S A,2007,104(6):1835 -1840.
[19]Baligand C,Gilson H,Ménard JC,et al.Functional assessment of skeletal muscle in intact mice lacking myostatin by concurrent NMR imaging and spectroscopy[J].Gene Ther,2010,17(3):328 -337.
[20]Gauthier MS,Couturier K,Charbonneau A,et al.Effects of introducing physical training in the course of a 16-week high-fat diet regimen on hepatic steatosis,adipose tissue fat accumulation,and plasma lipid profile[J].Int J Obes Relat Metab Disord,2004,28(8):1064-1071.
[21] Kemi OJ,Loennechen JP,Wisl?ff U,et al.Intensity-controlled treadmill running in mice:cardiac and skeletal muscle hypertrophy[J].J Appl Physiol,2002,93(4):1301 -1309.
[22]张 靓,唐朝枢.骨骼肌的内分泌功能[J].生理科学进展,2006,37(3):193 -198.
[23]Spiller MP,Kambadur R,Jeanplong F,et al.The myostatin gene is a downstream target gene of basic helix-loop-helix transcription factor MyoD[J].Mol Cell Biol,2002,22(20):7066 -7082.
Effects of Treadmill Running on Myostatin Expression in Soleus and Gastrocnemius Muscle of Rats
ZHANG Jing,LIU Xiaoyuan,YANG Hongtao
(School of P.E.and Sports,Beijing Normal University,Beijing 100875,China)
Objective:To observe the effects of treadmill running on the myostatin(MSTN)expression in soleus and gastrocnemius muscle of rats.Methods:Male 16-week-old SD rats were randomly divided into control group(n=10)and treadmill running group(n=10).The exercise rats underwent treadmill running at 26 m/min(60 min each,5 days/week for 8 weeks).The ratio of muscle weight/tibia length was measured,the cross section area of muscle was observed using the HE stain,and the mRNA and protein expression of MSTN were detected by real-time PCR and western bolt,respectively.Results:The soleus weight/tibia length and gastrocnemius weight/tibia length increased significantly in exercise group(both P<0.05)and the muscle cell cross section area enlarged markedly.Compared with the control group,treadmill training rats showed decreased MSTN mRNA and protein level in gastrocnemius muscle significantly(both P < 0.05),however,no difference were detected in soleus muscle.Conclusion:These results showed that the effects of treadmill running on MSTN expression were muscle fiber specific.
myostatin;muscle hypertrophy;soleus muscle;gastrocnemius muscle
G804.23
A
1004-0560(2012)05-0073-03
2012-04-12;
2012-05-19
中央高校基本科研业务费专项资金资助。
张 靓(1976-),女,副教授,博士,主要研究方向为运动与心血管活性物质研究;运动与骨骼肌活性物质研究。
责任编辑:乔艳春
猜你喜欢
体育科技文献通报(2022年3期)2022-05-23
中国畜牧杂志(2021年10期)2021-12-03
医学综述(2021年16期)2021-12-01
国际放射医学核医学杂志(2021年10期)2021-02-28
灌篮(2020年36期)2020-05-16
运动(2018年14期)2018-07-16
体育科学(2018年3期)2018-04-20
教育教学论坛(2018年1期)2018-01-18
中外医学研究(2017年15期)2017-06-29
心脑血管病防治(2015年3期)2016-01-15
