
禽流感抗原基因NA导入生菜的研究
2012-10-10 06:47范亚丽刘春林
湖南农业科学 2012年5期
范亚丽 ,李 进,阮 颖,刘春林
(1.阜阳师范学院生命科学学院,安徽 阜阳 236037;2.新疆建设兵团农一师十三团,
新疆 阿克苏 843302;3.湖南农业大学作物基因工程湖南省重点实验室,湖南 长沙 412328)
高致病力禽流感(High pathogenic avian fluenza,HPAI)是由A型流感病毒中的一些H5和H7亚型病毒引起的具有高度传染性、危害机体多种器官、高死亡率的禽类的一种系统性疾病,已被国际兽疫局列为A类烈性传染病[1-3]。自1905年禽流感病原被确认后,世界上共暴发了18次HPAI,其中8次是由H5亚型、10次是由H7亚型HPAIV引起的,给养禽业造成极大的经济损失[4]。禽流感病毒属正粘病毒科流感病毒属的A型流感病毒,基因组由8个分节段单股负链RNA组成,编码至少10种蛋白质[5]。第6节段RNA编码的神经氨酸酶(NA)是病毒囊膜表面的一种糖蛋白,其主要作用是水解细胞表面受体糖蛋白末端的N2乙酰基神经氨酸,以利于子代病毒的成熟和转移[6]。NA是诱导体液免疫的靶抗原之一,通过诱导机体产生特异性抗体,抑制病毒从感染细胞中释放,降低病毒在体内的复制,进而产生抗病毒作用[7]。用NA基因构建的DNA疫苗免疫动物,能诱导抗流感病毒致死攻击的免疫保护[8]。
生菜(Lactuca sativa var.capatata L.)为菊科莴属植物,是一种由国外引进的生食蔬菜,具有较高的营养价值,生长周期短,苣整株可以生食,再生和转化率较高,是一种用来表达外源蛋白的良好宿主植物。笔者采用农杆菌介导法将含有禽流感抗原基因NA的表达载体导入外植体生菜,初步研究了禽流感抗原基因NA在生菜基因组中的表达,旨在为以生菜作为植物生物反应器应用于大规模生产异源蛋白或疫苗提供科学依据。
1 材料与方法
1.1 材料
1.1.1 植物材料 生菜(Lactuca sativa var.capatata L.),品种:玻璃生菜(湖南农业大学作物基因工程实验室)。
1.1.2 菌株和载体 含目的DNA片段表达载体pBI121-NA的农杆菌菌株:Agrobacterium tumefaciens LBA4404。表达载体pBI121-NA由湖南农业大学作物基因工程实验室提供,李进[9]构建。表达载体pBI121-NA结构如图1所示。

图1 表达载体pBI121-NA结构
1.2 方法
1.2.1 无菌苗的获得 选取颗粒饱满的玻璃生菜种子,先用75%酒精浸泡30 s,再用40 mL 5%NaClO加入2滴吐温溶液浸泡15 min,无菌水冲洗3~5次,将种子均匀播种于1/2 MS培养基上,每个培养皿200粒,置于25℃光照培养,2~3 d获得培养一致的无菌苗。
1.2.2 农杆菌工程菌的准备 取携带pBI121-NA质粒的农杆菌LBA4404单菌落接种于3~5 mL含Kan、Str和Rif 3种抗生素YEB液体培养基中,于28℃、250 r/min振荡培养过夜;取培养液1 mL加入到含3种抗生素的50 mLYEB液体培养基中,28℃、250 r/min振荡培养4 h;测量菌液浓度OD600值,选取合适浓度的菌液4℃、5 000 r/min离心10 min,收集菌体,弃去上清液,用50 mL渗透液体培养基重悬菌体,备用。
1.2.3 子叶浸染转化 将生长2~3 d的生菜,利用子叶浸染转化法进行生菜的遗传转化。在重悬的菌液中加入0.05%β-巯基乙醇和100 mmol/L的AC混匀;然后用子叶放进OD600值为0.4~0.6菌液中浸染15 min,平放到共培养基上共培3 d;转入到诱导分化培养基上培养,两周后,将分化出的芽转移到苗生长培养基上,两周壮苗后,转入生根培养基;待苗长出发达的根系时,打开封口膜1 d,再将幼苗转入到蛭石固体培养基中炼苗一周;一星期后移栽到营养土中,移栽苗生长期间每3 d浇1次1/10 MS营养液,再经15 d左右移入大田。
1.2.4 转化植株的检测 (1)PCR扩增检测:根据目的片段的核苷酸序列设计引物,扩增481 bp的序列,引物序列如下。
NA_FB:5′-GAAGATCTTCATGAATCCAAATC AGAAGATAACAACCAT-3′
NA_RM:5′-CTCATTAATGTTCTGTGAGGGCT TCTG-3′
PCR反应体系为25 μL,反应条件:94℃预变性 4 min;94℃变性 40 s;60℃退火 40 s;72℃延伸120 s,共35个循环。72℃延伸7 min,16℃保存。
(2)RT-PCR检测:以未转基因的生菜叶片,转基因生菜叶片为材料,用RNA提取试剂盒进行生菜总RNA的提取,用Pronege DNAse试剂盒消化总RNA中含有的DNA,进行逆转录合成cDNA。分别以未转基因生菜叶片、转基因生菜叶片、未检测到目的片段的抗性生菜的反转录后cDNA、含目的片段DNA以及水为模板,以NA_FB,NA_FM为引物,进行PCR扩增。
2 结果与分析
2.1 卡那霉素和头孢霉素对生菜子叶分化能力的影响
生菜对卡那霉素(Kan)有一定的抗性,Kan对不定芽的诱导影响较大,明显表现出对生菜子叶分化的抑制作用,随Kan浓度升高,不定芽的诱导率以及平均出芽数逐渐降低。图2结果显示,当Kan浓度为80 mg/L时,不定芽的诱导率由对照的100%降低到85.4%,开始影响子叶的分化,平均出芽数由5.43个降低到2.85个;当Kan浓度达到100 mg/L时,不定芽的诱导和出芽数也大幅度减少;当Kan浓度为120 mg/L时,完全抑制生菜不定芽的诱导。因此,选择Kan 120 mg/L为诱导分化的筛选浓度。头孢霉素(Cef)对生菜植株的再生的抑制作用不是很明显,但仍然表现为随着抗生素浓度的提高,不定芽的诱导率逐渐降低,表明生菜对头孢霉素的抗性仍然能达到400 mg/L。因此,可以使用Cef作为农杆菌的生长抑制剂。
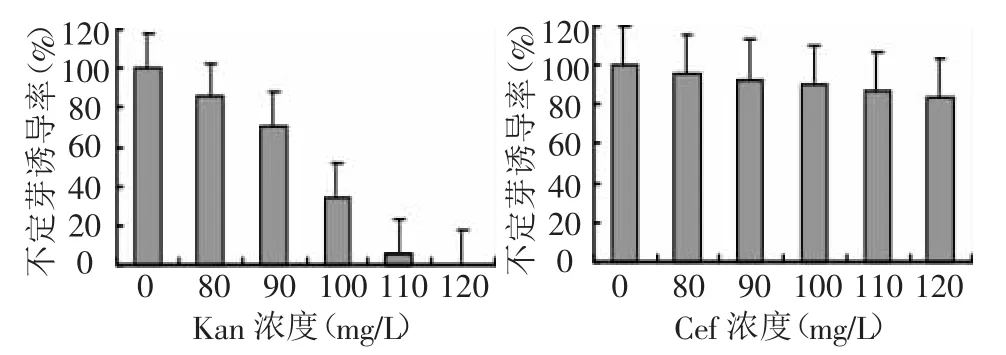
图2 抗生素对生菜子叶再生的影响
2.2 头孢霉素对生菜转化的影响
在农杆菌介导法转化生菜的过程中,抑菌是转化是否成功的一个关键因素[10]。图3结果表明:头孢霉素对生菜本身的选择压不明显,但随着抗生素浓度的提高,不定芽的诱导率逐渐降低,可选择头孢霉素抑制农杆菌生长达到试验目的。Cef浓度为50 mg/L时,抑菌率为58%;Cef浓度达到100 mg/L时,抑菌率为100%,当抑菌剂浓度更高时,抑菌率均达到100%。试验确定Cef的抑菌浓度为100 mg/L。
2.3 转基因生菜的检测
图3 头孢霉素对生菜转化的影响
2.3.1 抗性苗的PCR检测 把得到的抗性苗提取DNA作为模板,用2对筛选引物做PCR扩增,结果如图4所示:1~14泳道为提取DNA作为模板扩增结果,其中15泳道为对照(非转化植株),16泳道为质粒阳性对照,阳性植株的目的片段大小为481 bp,扩增带与预期大小相符。说明1、6、14泳道为转基因苗。图4中以不同转基因植株总DNA为模板进行PCR扩增,各带亮度不一,可能是受DNA提取纯度或基因在植物中的拷贝不同的影响。

图4 卡那霉素抗性苗的PCR检测
2.3.2 转基因生菜总RNA的提取 用试剂盒将总RNA抽提完成后,用紫外荧光光度计测定OD260和OD280,结果OD260/OD280为1.9,表明所提取的总RNA纯度较高。取10 μg RNA在无RNA酶的1.5%琼脂糖凝胶上进行电泳检测,结果如图5所示,1泳道为转基因生菜总RNA,2泳道为未转基因生菜总RNA。从图中可以看出1、2泳道中RNA样品没有降解,完全符合进行做反转录的要求。

图5 生菜总RNA的琼脂糖凝胶电泳
2.3.3 转基因生菜RT-PCR检测 分别以转基因生菜叶片、未转基因生菜叶片、未检测到目的片段的抗性生菜反转录后的cDNA、含目的片段DNA以及水为模板,经过PCR扩增得到的产物进行琼脂糖凝胶电泳(图6)。研究结果表明:5泳道为以含目的片段DNA为模板作阳性对照,其扩增产物大小为481 bp的条带,与marker相比较,目的片段位于300~500 bp之间,约481 bp,条带大小与预期的一致。1泳道为以转基因生菜反转录后的cDNA为模板,PCR扩增得到的产物为长约481 bp的特异性DNA片段,与阳性对照片段大小一致。2、3泳道分别为以未转基因生菜和未检测到目的片段的抗性生菜反转录后的cDNA为模板,4泳道以水为模板作阴性对照,均没有得到与阳性对照片段大小一致的条带。这说明目的片段只在转基因植株中检测到,而没有在未转基因植株中检测到目的片段,证明目的基因已经整合到生菜基因组中。

图6 转基因苗的RT-PCR检测
3 讨论
随着植物生物技术的发展,利用转基因植物生产外源蛋白或可食性疫苗已成为一个重要的研究和开发领域。植物生物反应器是指通过基因工程途径,以常见的农作物作为“细胞工厂”,通过大规模种植生产具有高附加值的医用蛋白、疫苗、工农业用酶、特殊碳水化合物等植物反应器[11]。其最大的优点是可以大规模、廉价、安全地生产动物蛋白或疫苗[12],特别是用于免疫和治疗性疫苗的生产,已随着研究的深入越来越显示出其独特的优势[13]。基因工程领域的研究进展,使得植物体正在成为具有重要经济价值的异源蛋白(包括药用蛋白)的生产体系。据不完全统计,迄今为止,国外已经有几十种药用蛋白质或多肽在植物中得到成功表达,其中包括了人的细胞因子、表皮生长因子、促红细胞生成素、干扰素、生长激素、单克隆抗体和可作为疫苗用的抗原蛋白等[14]。
笔者通过农杆菌介导法转化生菜,已经获得卡那霉素抗性苗105株,用特异引物对抗性植株进行PCR检测,其中6株为转基因植株。经RT-PCR检测可知,禽流感抗原基因NA已经在生菜基因组中得到整合并初步验证在转录水平上得以表达。通过阳性株的无性繁殖可以扩大阳性植株的数量,为禽流感抗原基因NA在生菜中的表达、编码蛋白免疫原性以及能否在后代中稳定遗传的研究打下基础。
[1]黄庆华,张果平,王 可.禽流感病毒NP蛋白单克隆抗体的研制和鉴定[J].江西农业学报,2010,22(12):141-143.
[2]于 博,章振华,姜北宇,等.一株H9N2亚型禽流感病毒全基因组序列的遗传变异分析 [J].安徽农业科学,2010,38(1):507-509,526.
[3]Shen J,Zhang Z H,Jiang B Y,et al.Complete genome sequencing and genetic variation analysis of two H9N2 subtype avian influenza virus strains[J].Agricultural Science&Technology,2011,12(2):291-294.
[4]Tang X Y,Tian G B,Zhao C S,et al.Isolationand characterization of prevalent strains of avian influenza viruses in China[J].Chinese Journal Animal Poultry Infection Disease,l998,20(1):l-5.
[5]Alexander D J.A review of Avian Influenza in Different Bird Speices[J].Veterinary Micorbiology,2000,74:3-13.
[6]Palese P ,Tobita K,Ueda M ,et al.Characterization ofTemperature Sensitive Influenza Virus Mutants Defective in Neuraminidae[J].V i rology,1974,61:397-410.
[7]Webster R G,Reay P A ,Laver W G.Protection against Lethal Influenza with Neuraminidase[J].Virology,1988,164:230-237.
[8]Sandbulte M R ,J imenez G S ,Boon A C M ,et al.Cross-Reactive Neuraminidase Antibodies Afford Partial Protection against H5N1 in Mice and Are Present in Unexposed Humans[J].PLOS Medicine,2007,4:265-271.
[9]李 进,阮 颖,龚 剑,等.禽流感抗原基因NA,HA的克隆及其表达载体的构建[J].现代生物医学进展,2008,8(3):449-452.
[10]莽克强.农业生物工程[M].北京:化学工业出版社,1998.
[11]Oersbo M,Okkels F T.A novel principle for the selection of transgenic plant cells:positive selection[J].Plant Cell Reports,1996,16:219-221.
[13]Hiatt A,Cafferkey R,Bowdish B.Production of antibodies in transgenetic plants[J].Nature,1989,342:76-78.
[13]Tacket C O,Mason H C,Losonsky G,et al.Immunogenicity in humans of a recombinant bacterial antigen delivered in a transgenic potato[J].Nature Medicine,1998,5(4):607-609.
[14]Mason H S,Ball J M,Shi J J,et al.Expression of hepatitis B surface antigen in transgenic plants[J].Proc Natl Acad Sci USA,1992,89:11745-11749.
猜你喜欢
中老年保健(2022年6期)2022-08-19
数学大王·低年级(2020年8期)2020-08-14
农家参谋(2020年5期)2020-06-15
生物工程学报(2019年1期)2019-01-30
启蒙(3-7岁)(2017年4期)2017-06-15
食品与健康(2017年3期)2017-03-15
美食堂(2017年2期)2017-02-14
小雪花·初中高分作文(2016年9期)2016-05-14
当代畜禽养殖业(2014年7期)2014-02-27
当代畜禽养殖业(2014年6期)2014-02-27
