
不同生境中飞机草的群落结构及其生物多样性
2012-09-18 06:33:01蒲高忠唐赛春潘玉梅韦春强刘明超
草业科学 2012年3期
蒲高忠,唐赛春,潘玉梅,韦春强,刘明超,2
(1.广西壮族自治区、中国科学院广西植物研究所,广西桂林541006;2.广西师范大学生命科学学院,广西桂林541004)
不同生境中飞机草的群落结构及其生物多样性
蒲高忠1,唐赛春1,潘玉梅1,韦春强1,刘明超1,2
(1.广西壮族自治区、中国科学院广西植物研究所,广西桂林541006;2.广西师范大学生命科学学院,广西桂林541004)
采用样方法,研究了桂西南喀斯特地区外来入侵植物飞机草(Eupatoriumodoratum)对人工疏林、弃耕地和荒山灌丛的群落组成、结构特征以及植物物种丰富度的影响。结果表明,3种生境中共有植物79种,隶属45科50属,群落灌层和草层分层不明显;但植物种类、数量和垂直结构在3种生境中存在差异;影响飞机草群落分布组成的主要因素是人为干扰和光照,其贡献率分别为42.88%和24.17%,据此将调查样点分为3个聚类群;当飞机草重要值大于0.51时,样方物种丰富度随重要值的增加而减小,二者之间具有显著相关性。表明飞机草的入侵对生物多样性有不利的影响,而这种影响又受人为干扰强度的限制。
飞机草;群落结构;主成分分析;生物多样性
外来有害物种导致的生物入侵在全球范围内不断加剧,被认为是21世纪最棘手的环境问题之一。评价外来植物入侵对其入侵地生物多样性与生态系统功能的影响已成为入侵生态学研究的热点,其中群落或生态系统的可入侵性又是入侵生态学最重要的一个方面[1-3]。植物入侵常会造成入侵地植物区系大范围的均化[4],即由于大量的植物入侵造成的植物地理单元由不均一转变为相似的地理单元,从而导致入侵地生物多样性丧失[5-10]。
飞机草又名香泽兰,菊科泽兰属丛生型多年生草本或亚灌木,原产南美洲安第斯山。20世纪30年代由泰国传入我国云南地区,此后迅速蔓延、传播,现已传至贵州、广西、海南、广东、台湾等省(区),侵占耕地和林地,对农、林、牧业及自然保护区的生物多样性保护等带来严重危害,对入侵地造成巨大生态和经济损失[11-13]。飞机草进入我国已有70多年的历史,人们对其的研究主要集中在飞机草的生物学特性[14-16]、传播途径[12]、入侵机制[17-21]、控制策略和开发利用等方面[22-24],群落分布研究显示,飞机草群落的种类组成、结构与外貌特征等随着分布区域、生境和人为干扰等因素的差异而有所不同,但这种差异在生态功能脆弱的喀斯特地区是否存在以及差异程度的大小目前并不清楚[12-13,25-26]。因此,本研究选取广西喀斯特地区3种不同人为干扰生境中的飞机草群落样地为研究对象,调查不同生境中飞机草群落的种类组成及其结构特点,并通过比较分析飞机草的入侵对生物多样性的影响,以期为飞机草的预警和治理提供科学依据。
1 研究方法
1.1 研究地自然概况 研究区位于桂西南喀斯特区域,3种生境分别为位于广西龙州县的人工疏林、金龙乡的弃耕地和平果县龙河地区的荒坡灌丛,分别标为样地A、B、C。样地A属于峰林谷地,含较多棕色石灰土,山上为4~5龄砍头树人工疏林,人为干扰程度最强,位于22°36′50″N,106°47′44″E,海拔184m;样地B处于低山丘陵地带,石灰土含量多,生于公路旁土山弃耕地(约4年),人为干扰程度较弱,位于22°33′20″N,106°40′42″E,海拔274m;样地C属典型的石漠化区,位于23°22′30″~23°24′00″N,107°23′38″~107°25′30″E,海拔110~570m,该地突出特点为岩石裸露、土壤浅薄、植被覆盖率不足10%,石漠化严重,土地生产力较低[27],但人为干扰程度最弱。
1.2 群落调查 研究在2006年和2007年每年7月中、下旬进行。采用样方法,对上述不同生境下飞机草植物群落内的植物种类和分布情况进行调查,在3个样地选定10个样点(样地A、B各2个,样地C 6个)。每个样点设置4个样方(2m×2m)。调查样方中所有杂草的种类、数量、盖度,并观察各样地的光照条件、土壤水分条件等环境因素。每种杂草的相对多度=每种杂草数目/样方内所有杂草总数;每种杂草的相对频度=每个样点中出现该种杂草的样方数/总样方数;每种杂草的重要值=(该种杂草的相对多度+该种杂草的相对盖度+该种杂草的相对频度)/3。调查地点及样点的生境描述见表1。
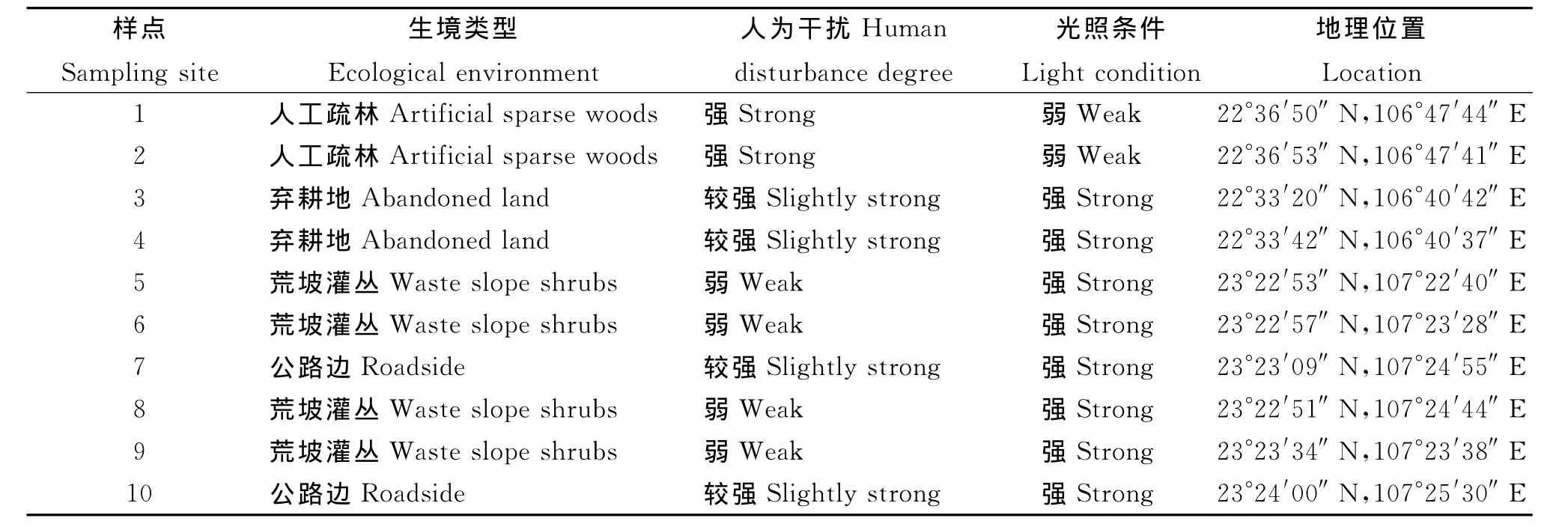
表1 10个样点的生境类型Table 1 Habitat types of sampling sites
1.3 数据处理 选取发生频率(发生频率=出现某种杂草的样方数/样方总数)>10%的36种植物重要值作为分析指标,10个样点作为分析单位,构成2个原始数据矩阵。应用SPSS 17.0统计软件,对原始数据进行主成分分析。主成分分值通过标准化转换得到,正号表示它对主成分的影响为正相关的关系,负号反之。此外,对分析结果进行生态学意义上的解释,研究不同生境条件下飞机草的分布及其对生物多样性的影响。
2 结果
2.1 群落种类组成比较 各样地群落组成表明,3种生境中共有植物79种,隶属45科50属。其中样地A群落中植物种类最少,主要由砍头树和草本植物组成,共计18种,隶属12科18属,其中菊科(4种)和禾本科(3种)植物居多;样地B分属草本群落,群落中共计植物28种,隶属21科28属,其中菊科外来入侵植物多达6种,禾本科和唇形科各2种,其余均为1种;样地C群落成分比较复杂,主要由木本、藤本植物组成,间杂少量草本,共35种,分属22科34属,禾本科5种,大戟科和豆科各为4种,菊科3种,防己科、桑科和鼠李科各2种,其余各1种。在各群落中,除上述优势科外,其余大多为单科、单属。表明随着人为干扰程度由弱至强,飞机草群落向着科、属和种组成更为单一、简单的方向发展。
2.2 群落结构特征 群落的结构是指群落的成层现象。3个样地飞机草群落结构特征的研究结果(表2)显示,乔木层仅在样地A存在;灌、草层则在3个样地中均存在,并且结构简单,分层不明显,总体高度0.5~2.5m。样地A由于人为干扰严重,乔木层仅砍头树1种,高4~9m;灌木层主要由飞机草组成,而草本层植物种类和数量明显增加。样地B为撂荒4年以上的弃耕地,所形成的群落高度仅0.3~1.8m。分灌木、草层2层,灌木层中飞机草和小飞蓬的数量占该层总株数的80%以上,其间散生着雷公根、八角枫(Alangiumchinense)、五节芒等;草本层则散生于其中。样地C为典型的石漠化区的荒坡灌丛,与样地B相同,也仅具灌木、草本层。但由于该样地受人为活动影响较小,其灌木层优势明显,高0.8~2.0m;草本层除了飞机草、黄荆条、雀梅藤、柞木、潺槁树等植物幼苗外,整个样地散生有少量水蔗草、千里光、薄荷等。
2.3 主成分分析 第1主成分贡献率为42.88%,第2主成分贡献率为24.17%(表3)。从前2个主成分负荷量来看,第1主成分特征向量呈现较大正相关的植物有小飞蓬、荩草、艾蒿、雷公根等,多生于人工疏林、弃耕地以及公路边等人为干扰程度较强的生境中;呈现较大负相关的植物有红背山麻杆、野牡丹、盐肤木、潺槁树、勾儿茶等,这些植物生长在人为干扰程度较弱的石山灌丛生境中。由此不难看出,第1主成分轴所代表的生态学意义是人为干扰程度,且干扰程度越高,第1主成分特征向量值越小。从第2主成分轴特征向量值来看,呈现较大正相关的是金腰箭、海金沙、茜草、飞扬草和地桃花等,这些杂草多生长在人工疏林下或较为阴湿和遮阴的草本层中,光照强度较弱;呈现较大负相关的杂草有砍头树、黄荆条、飞龙掌血等,它们喜欢阳光较为充足的荒山灌丛生境中。可见,第2主成分轴所代表的生态学意义是光照强度。综上所述可知,飞机草更倾向于生长在光照较强和人为干扰程度较弱或中度干扰的生境中,影响飞机草生长分布区与危害的主要因素是人为干扰程度和光照强度。
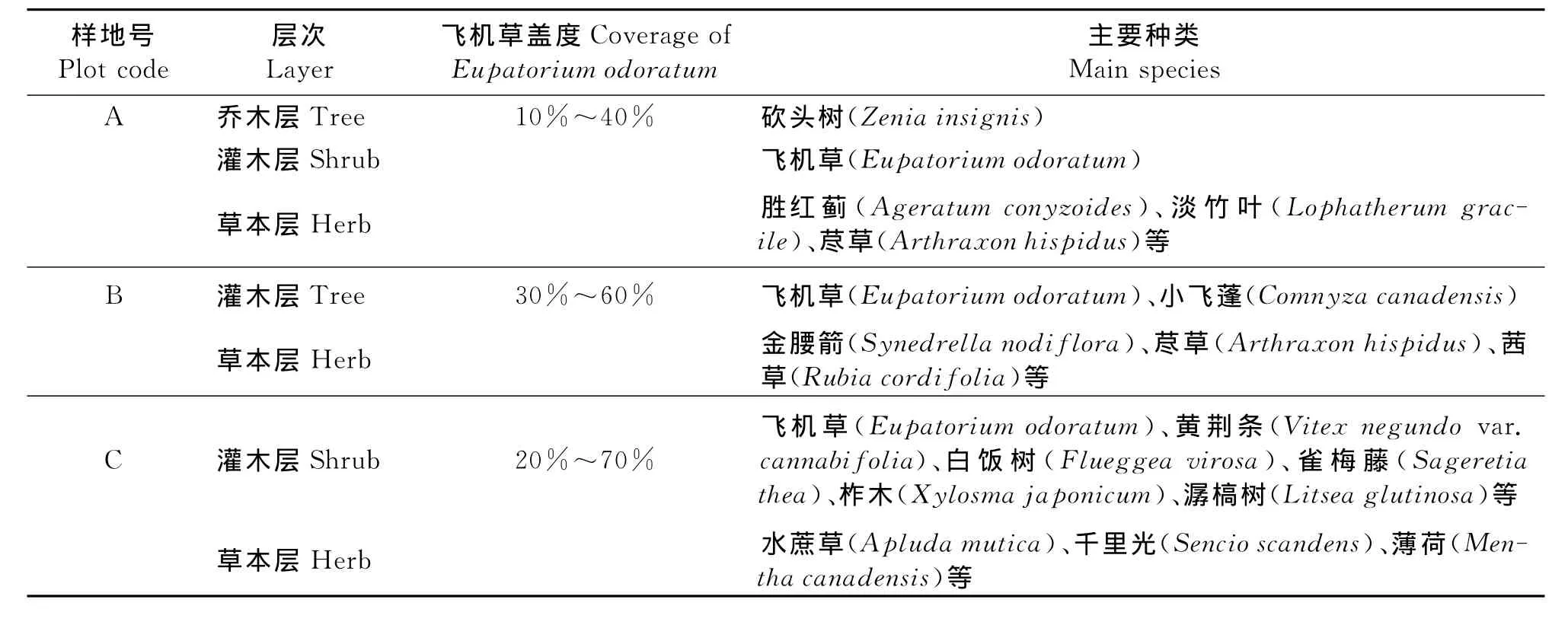
表2 不同生境飞机草群落的结构特征Table 2 Community structure characteristics ofEupatoriumodoratumunder different habitats
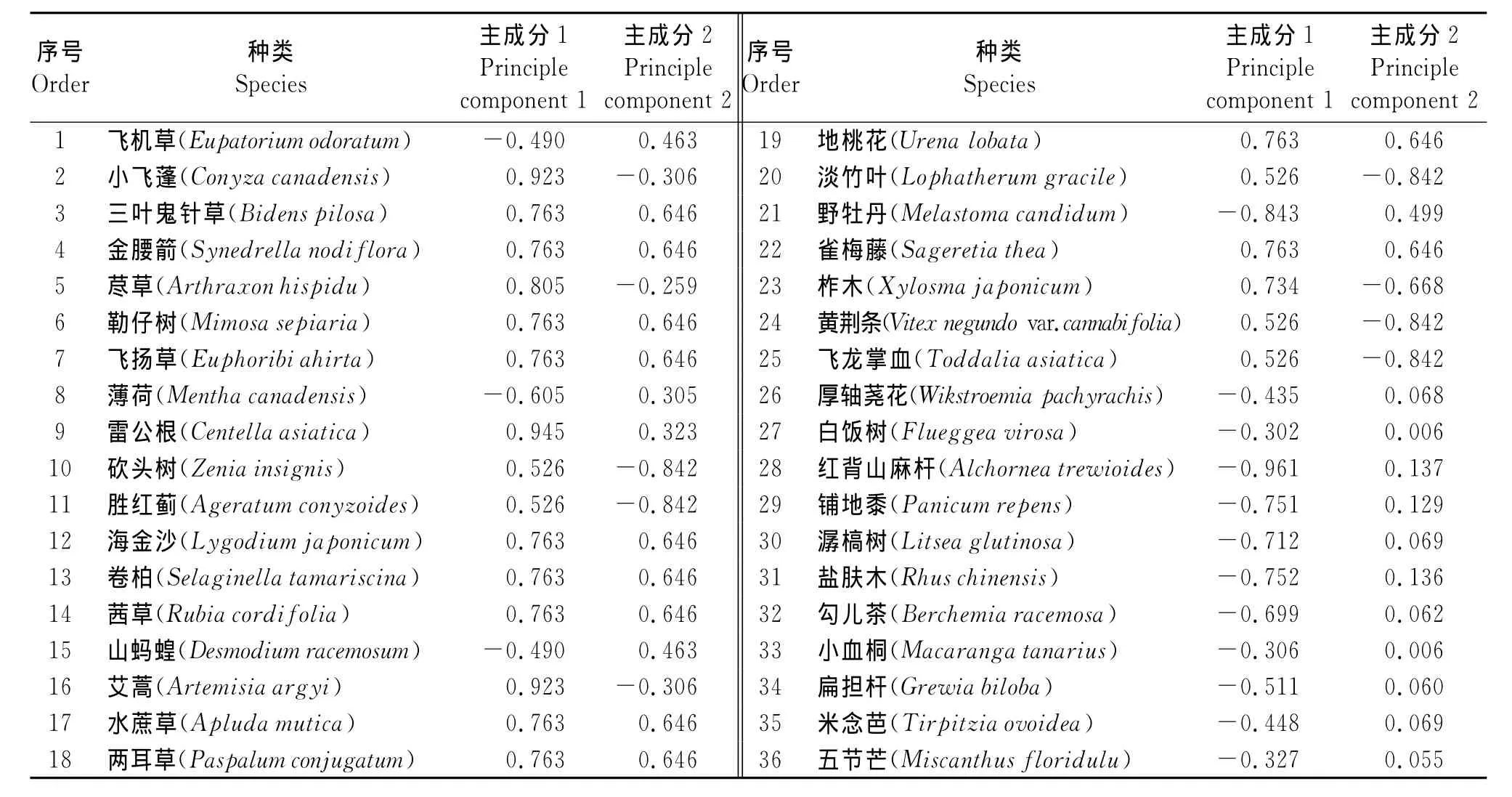
表3 36种植物的前两个主成分负荷量Table 3 Contribution of first two principle components of the 36plant species
2.4 不同生境对飞机草种群密度的影响 对 3种生境下飞机草分布的10个样点进行Q型聚类分析(图1),结果显示,有飞机草分布的样点可分为聚类群Ⅰ、Ⅱ和Ⅲ。聚类群Ⅰ主要分布于人工疏林,人为干扰程度强,但由于被乔木(砍头树)遮挡,飞机草的光照相对不足,因而种群密度较小,表明在高强度人为干扰情况下,飞机草的生长和扩散趋势受到一定的限制。聚类群Ⅱ分布在公路旁或弃耕地等人为干扰较强的开阔地带,光照充足,土壤含水量较高,故种群密度较大,并以飞机草、小飞蓬、金腰箭等为主。聚类群Ⅲ包括的样点较多,均分布在人为干扰程度较弱的石山灌丛中,光照充足,种群密度最大,说明飞机草适合生长于阳光充足的生境中,并且由于它强大的扩散对策优势而占据了其他物种的生存空间。可见,对于飞机草群落进行高强度的人为干扰(主要种植本地生长迅速的乔木等),可有效降低飞机草的密度和生存空间。
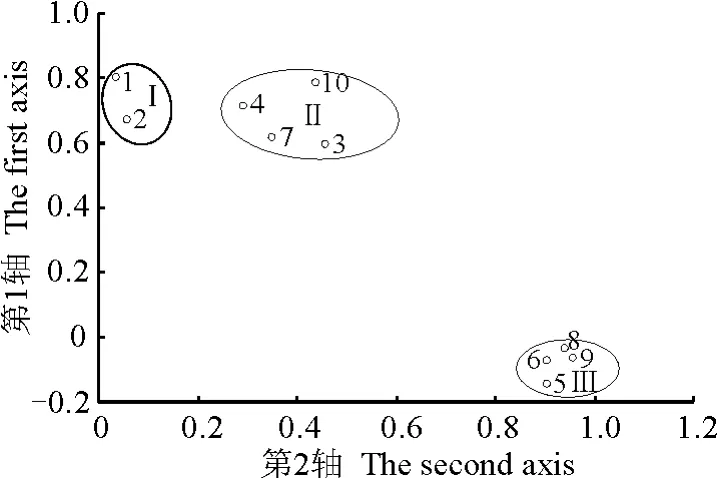
图1 10个样点的三维排序散点图Fig.1 3-D scatter plot of ten sampling site
2.5 不同生境下飞机草入侵对生物多样性的影响 飞机草在3种生境中均有分布,即相对频度相同(表4)。受人为活动干扰最强的样地A中相对盖度最低,且与其他样地间均存在显著差异(P<0.05),而在受人为活动干扰较弱的样地B和C之间差异不显著(P>0.05)。同时,样地A中飞机草个体数量也最少,相对密度远小于其他样地,并在3个样地间均存在显著差异。随着人为干扰程度(样地A>样地B>样地C)的减弱,飞机草重要值呈增大趋势,并在各样地之间存在显著差异。其中在样地C植物群落中的重要值最大,样地A的最小,样地B中的则介于二者之间(表4)。表明样地A生境群落中的优势最小,人工种植的砍头树不仅干扰了飞机草的生长,也干扰了其他杂草的生长。
表4 不同生境飞机草的分布特点与群落多样性
Table 4 Distribution characteristics ofEupatoriumodoratumcommunity and biodiversity at different habitats
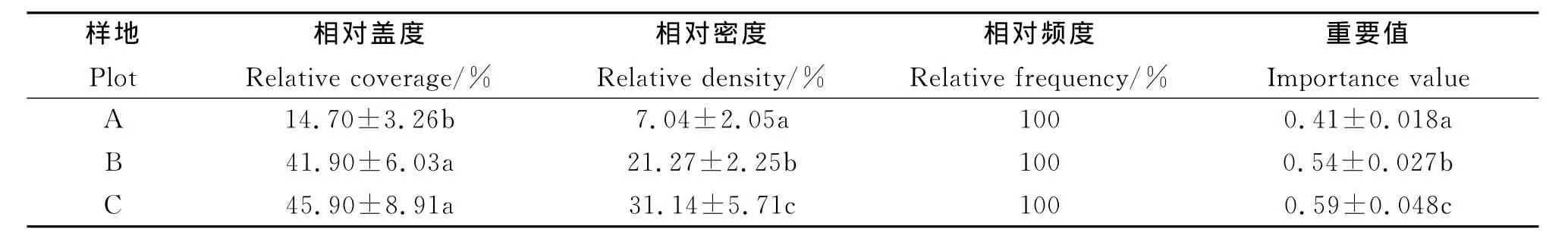
注:同列不同小写字母表示显著差异(P<0.05)。Note:Different lower case letters within the same column show significant diffrence at 0.05level.
将各样方飞机草重要值划分为5个梯度:0.35~0.45,0.45~0.53,0.53~0.55,0.55~0.60,0.60~0.65。求得飞机草各梯度平均重要值及对应样方物种平均丰富度,并对二者进行曲线模拟。二者拟合曲线表达式为y=1 181.9x2+1 189.3x-279.3(R2=0.922 1),具有显著相关性。当飞机草重要值较低时,样方物种丰富度随重要值得增加而增加;当重要值大于0.51时,样方物种丰富度随重要值的增大而下降(图2)。
上述结果表明,在不同生境中,飞机草对群落中生物多样性有显著的影响,同时,这种影响又一定程度上受人为干扰强度的限制。
3 讨论与结论

图2 飞机草的发生数量对植物物种丰富度的影响Fig.2 Effects ofEupatoriumodoratumnumber on plant richness
飞机草群落的种类组成、结构与外貌特征等随着分布区域、生境、发育阶段和人为干扰等因素的差异而有所不同[9]。对各入侵地生境物种组成研究分析表明,受强烈干扰的样地A群落中植物种类最少,受干扰程度较低的样地B次之,而在受干扰程度最弱的样地C则最多。可见,由于受生境和干扰程度等因素的影响,广西喀斯特地区飞机草群落在不同生境之间存在较大的差异,即随着人为干扰程度由弱至强,飞机草群落向着科、属、种组成等更为单一、简单的方向发展,其结构特征也由灌木层优势明显向不明显发展。
外来物种的成功入侵除受自身的生物学特性制约外,还与入侵地的气候特点、生境类型密切相关,如果入侵区域的环境条件与原产地类似,那么外来种成功入侵的机率就会上升,反之,其生长就会受到一定的限制[11,24]。主成分分析表明,人为干扰程度以及光照条件是决定飞机草群落组成的两个重要因素。飞机草适合生长于阳光充足的生境中,并由于它强大的扩散优势而占据了其他物种的生存空间。同时,随着人为干扰程度的增强,飞机草的重要值(发生率)反而有所下降,即低干扰程度生境中的重要值高于高干扰程度的生境。这主要得益于5年龄的砍头树人工疏林有效地降低了机草的光照强度,且飞机草喜光而不耐阴[12],所以,在此生境下其生长和扩散受到一定的限制。对飞机草群落进行高强度的人为干扰(主要种植本地生长迅速的乔木等),可有效降低飞机草的密度和生存空间。此外,对飞机草各梯度平均重要值及样方物种平均丰富度进行曲线模拟的结果表明,当样方中飞机草重要值>0.51时,物种丰富度开始下降,二者存在显著相关性。表明在不同生境中,飞机草对群落中的生物多样性有显著影响,同时,这种影响又一定程度上受人为干扰强度的限制。
[1]陈慧丽,李玉娟,李博,等.外来植物入侵对土壤生物多样性和生态系统过程的影响[J].生物多样性,2005,13(6):555-565.
[2]D’Antonio C M,Kark S.Impacts and extent of biotic invasions in terrestrial ecosystems[J].Trends in Ecology and Evolution,2002,17:202-204.
[3]许凯扬,叶万辉,曹洪麟,等.植物群落的生物多样性及其可入侵性关系的实验研究[J].植物生态学报,2004,28(3):385-391.
[4]Hejda M,Pyšek P,Jarošik V.Impact of invasive plants on the species richness,diversity and composition of invaded communities[J].Journal of Ecology,2009,97: 393-403.
[5]徐汝梅,叶万辉.生物入侵——理论与实践[M].北京:科学出版社,2003:15.
[6]Rejmánek M,Richardson D M,Higgins S I,etal.Ecology of invasive plants:state of the art[A].In:Mooney H A,Mack R N,McNeely J A,etal.Invasive Alien Species:A New Synthesis[M].Washington D C:Island Press,2005:104-161.
[7]Schwartz M V,Thorn J H,Viers J H.Biotic homogenization of the California flora in urban and urbanizing regions[J].Biological Conservation,2006,127:282-291.
[8]Kühn I,Klotz S.Urbanization and homogenization:Comparing the floras of urban and rural areas in Germany[J].Biological Conservation,2006,127:292-300.
[9]张震,徐丽,朱晓敏.喜旱莲子草对不同生境植物群落多样性的影响[J].草业学报,2010,19(4):10-15.
[10]郭连金,徐卫红,孙海玲,等.空心莲子草入侵对乡土植物群落组成及植物多样性的影响[J].草业科学,2009,26(7):137-142.
[11]刘金海,黄必志,罗富成.飞机草的危害及防治措施简介[J].草业科学,2006,23(10):73-77.
[12]余香琴,冯玉龙,李巧明.外来入侵植物飞机草的研究进展与展望[J].植物生态学报,2010,34(5):591-600.
[13]全国明,章家恩,徐华勤,等.外来入侵植物飞机草的生物学特性及控制策略[J].中国农学通报,2009,25(9):236-243.
[14]奎嘉祥,匡崇义,和占星,等.中国云南南部建植臂形草混播草场防治飞机草的研究[J].中国草地,1997(5):55-58.
[15]曹洪麟,葛学军,叶万辉.外来种飞机草在广东的分布与危害[J].广东林业科技,2004,20(2):57-59.
[16]Ambika S R.Ecological adaptations ofChromolaena odorata(L.)King and Robinson[A].In:Ferrar P,Muniappan R,Jayanth K P.Proceedings of the Fourth International Workshop on Biological Control and Management of Chromolaena odorata[C].India:Bangalore,1996.
[17]Bais H P,Vepachedu R,Gilroy S,etal.Allelopathy and exotic plant invasion:From molecules and genes to species interactions[J].Science,2003,301:1377-1380.
[18]Feng Y L,Lei Y B,Wang R F,etal.Evolutionary tradeoffs for nitrogen allocation to photosynthesis versus cell walls in an invasive plant[J].Proceedings of the National Academy of Sciences of the United States of America,2009,106:1853-1856.
[19]张常隆,李扬苹,冯玉龙,等.表型可塑性和局域适应在紫茎泽兰入侵不同海拔生境中的作用[J].生态学报,2009,29(4):1940-1946.
[20]Sangakkara U R,Attanayake K B,Dissanayake U,et al.Allelopathic impact ofChromolaenaodorata(L.)King and Robinson on germination and growth of selected tropical crops[J].Journal of Plant Diseases and Protection,2008,21:321-326.
[21]潘玉梅,唐赛春,蒲高忠,等.岩溶区土著植物黄荆条和红背山麻杆水浸提液对入侵植物飞机草萌发的影响[J].中国岩溶,2008,27(2):97-102.
[22]Bamba D,Bessiere J M,Marion C,etal.Essential oil ofEupatoriumodoratum[J].Planta Medica,1993,59(2):184-185.
[23]付玉杰,刘威,侯春莲,等.RP-HPLC/二极管阵列检测器同时测定飞机草中3种黄酮[J].应用化学,2007,24(12):1452-1454.
[24]刘晓妹,蒲金基,蒙美英.飞机草不同溶剂粗提液抑菌活性的测定[J].广西热带农业,2004(6):1-3.
[25]李志刚,郑启恩,黎桦,等.广西隆安屏山石灰岩山地飞机草群落特征分析[J].热带亚热带植物学报,2006,14(3):196-201.
[26]吴邦兴.滇南飞机草群落的初步研究[J].西华师范大学学报(自然科学版),1981(1):40-64.
[27]韦兰英,袁维圆,尤业明,等.岩溶石漠化区牧草植物地上部分生物量的动态变化[J].草业科学,2009,26(10):73-79.
Eupatorium odoratumcommunity structure and its effects on species richness of native vegetation community under different habitats in Karst Area of Guangxi
PU Gao-zhong1,TANG Sai-chun1,PAN Yu-mei1,WEI Chun-qiang1,LIU Ming-chao1,2
(1.Guangxi Institute of Botany,Guangxi Zhuang Autonomous Region and the Chinese Academy of Sciences,Guilin 541006,China;2.College of Life Science,Guangxi Normal University,Guilin 541004,China)
Eupatoriumodoratum,which is a worldwide perennial invasive alien weed,usually spreads rapidly,and poses a serious threat to crops,forest,native vegetation and biodiversity.In order to understand characteristics ofChuomolaenaodoratacommunity structure in different habitats and its effects on plant species richness of native vegetation community,threeC.odoratacommunities from different habitats were chosen for establishing three sample plots(plot A,B and C)in Longzhou county and Pingguo county of Guangxi.Plant species composition,structure characteristics,distributions ofC.odorataand its effects on plant species richness of native vegetation in each plot were investigated and analyzed.The results showed that there were a total of 79species,which belonged to 50genera of 45families,under the three habitats.The community structures were simple,including there layers(tree,shrub and herb)in plot A,two layers(shrub and herb)in plot B and C,but the plant species,quantity and vertical structure were different.Through the principle component analysis of 36species with occurring frequency>10%,we found that the human disturbance and light condition were the key factors affected theC.odoratacommunity structure,which contributed 42.88%and 24.17%,respectively.According to the two factors,all the samples could be divided into three cluster groups.The analysis of relationship between the importance value ofC.odorataand plant species richness of the communities indicated that when the former reached to 0.51,the latter decreased with the rise of the former,which demonstrated a significant correlation between them.This study suggests that the invasion ofC.odoratahas an adverse effect on biodiversity,but the effect depends on the intensity of human disturbance.
Eupatoriumodoratum;community structure;principal component analysis;biodiversity
TANG Sai-chun E-mail:tangs@gxib.cn
S812;Q948.15
A
1001-0629(2012)03-0447-06
2011-05-03 接受日期:2011-07-17
广西科学研究与技术开发计划项目(桂科攻0992026-5、0719005);广西自然科学基金项目(2011GXNSFE018005);广西植物研究所科学研究基金(桂植业10011);国土资源部、广西壮族自治区岩溶动力学重点实验室开放课题(KDL2010-03)
蒲高忠(1980-),男,甘肃西和人,助研,硕士,主要从事外来入侵植物研究。E-mail:gzpu@gxib.cn
唐赛春 E-mail:tangs@gxib.cn
猜你喜欢
湖北植保(2022年4期)2022-08-23 10:51:52
林业勘查设计(2022年1期)2022-02-15 05:35:12
乡村科技(2021年17期)2021-10-20 08:50:56
安顺学院学报(2021年4期)2021-09-16 08:11:08
土壤(2021年1期)2021-03-23 07:29:06
林业调查规划(2020年3期)2020-06-03 07:02:36
摄影之友(影像视觉)(2019年2期)2019-03-05 08:27:20
土壤学报(2017年5期)2017-11-01 09:21:27
中国篆刻(2017年5期)2017-07-18 11:09:30
诗歌月刊(2014年1期)2014-03-11 17:26:03
