
几株益生乳酸菌对Caco-2细胞的黏附及其对致病菌黏附的影响
2012-09-11 07:35:20李卫芬崔志文余东游
动物营养学报 2012年10期
白 洁 李卫芬 黄 琴 崔志文 余东游 黄 怡,2*
(1.浙江大学动物科学学院,浙江大学动物分子营养学教育部重点实验室,杭州 310058;2.广西大学动物科学技术学院,南宁 530005)
黏附是微生物与宿主肠道相互作用的第1步,也是进入肠道的微生物发挥作用的前提[1]。黏附和定植于宿主肠道的能力是益生菌的评价标准之一,益生菌株必须对上皮细胞组织具有黏附性才能长期在肠道中存活并发挥其作用[2]。黏附在肠道的益生菌可以通过空间占位,产生抑菌物质等对肠道病原菌发挥抑制作用,调整肠道菌群平衡。同时,黏附也是感染性细菌致病的第1步,如果能抑制病原菌对宿主细胞的黏附,往往能抑制病原菌的致病作用[3]。因此,人们常将益生菌的黏附性能及其对致病菌的黏附抑制作用作为菌株质量评定的重要指标。目前,国内外有关乳酸菌对细胞的黏附及其对病原菌的黏附抑制作用的研究报道较多,并且发现一些有较好黏附性能的益生乳酸菌[4-5],但是关于其对肠上皮细胞的黏附是否具有菌属和宿主特异性方面的研究报道较少。肠上皮样细胞系Caco-2细胞是从人结肠腺癌中建立的,是目前研究细菌或病毒与肠道黏膜黏附应用最广泛的细胞模型。该细胞模型能够在体外进行形态功能分化,表现出成熟肠上皮细胞的特性。因此,本试验采用目前国内外学者普遍使用的体外培养Caco-2细胞的方法来评价几株不同来源的益生乳酸菌的黏附能力及其对大肠杆菌K88和鼠伤寒沙门氏菌的黏附抑制作用,并探讨不同种属及来源的菌株对Caco-2细胞的黏附性能的差异,为乳酸菌更好地应用于生产及探讨其黏附机制提供初步的理论依据。
1 材料与方法
1.1 菌株和细胞株
L1~L6 6个乳酸菌菌株,L1:鼠李糖乳杆菌(Lactobacillus rhamnosus,编号为 6001),购自中国工业微生物菌种保藏管理中心;L2:食果糖乳杆菌(Lactobacillus fructivorans,编号为6052)冻干粉,购于中国食品发酵工业研究院;L3:乳杆菌(Lactobacillus),分离于健康鸡肠道;L4:粪肠球菌(Enterococcus faecalis,编号为20426)冻干粉,购于中国食品发酵工业研究院;L5:屎肠球菌(Enterococcus faecium),由浙江大学饲料科学研究所微生物与基因工程实验室保存;L6:乳酸乳球菌(Lactococcus lactis,编号为23609),购自中国工业微生物菌种保藏管理中心。2个致病菌菌株,K88:大肠杆菌(Escherichia coli)K88,购自国家兽医微生物菌种保藏管理中心;S:鼠伤寒沙门氏菌(Salmonella typhimurium),购自国家兽医微生物菌种保藏管理中心。Caco-2细胞购自中国科学院上海生命科学研究院细胞资源中心。
1.2 主要试剂和设备
MRS、GM17和 LB培养基(英国 Oxoid公司);DMEM培养液、0.25%胰酶[含0.02%乙二胺四乙酸(EDTA)]、D-hank’s平衡盐溶液(美国Gibco公司);胎牛血清(杭州四季青生物工程材料有限公司);青霉素和链霉素(美国Sigma公司);Triton X-100和台盼蓝染色液(美国Ameresco公司)。细胞培养瓶及培养板(美国Gibco公司);5804R高速冷冻离心机(德国Eppendorf公司);HERA cell 150 CO2细胞培养箱(美国Thermo公司);CX41-12C02倒置显微镜(日本Olympus公司);NOVEL XSZ-N107CCD光学显微镜(宁波永新光学股份有限公司);UV-2100紫外可见分光光度计(尤尼柯上海仪器有限公司);Synergy 4酶标仪(美国BioTek公司)。
1.3 试验方法
1.3.1 供试菌液制备
除了乳酸乳球菌用GM17培养基外,其他乳酸菌都用MRS培养基培养。乳酸菌、大肠杆菌K88和鼠伤寒沙门氏菌-80℃保存,试验前乳酸菌接种于新鲜配制的GM17或MRS液体培养基,30℃静止培养24~36 h,并转接2代。大肠杆菌K88和鼠伤寒沙门氏菌接种于LB培养基,37℃摇床培养过夜,并转接2代。分别将生长至对数生长期的乳酸菌(16 h)、大肠杆菌K88(8 h)和鼠伤寒沙门氏菌(10 h)在4 000 r/min、15℃离心10 min,收集菌体,用无菌磷酸盐缓冲液(PBS)洗涤2次,最后重悬于无菌PBS中,调节细菌的浓度为2×108CFU/mL。
1.3.2 细菌荧光标记
按照文献[6]和[7]的方法标记细菌。把上述准备好的乳酸菌、大肠杆菌K88和鼠伤寒沙门氏菌(1×109CFU/mL)悬浮于100 mg/mL异硫氰酸荧光素(FITC)中,37℃暗处作用1 h,用PBS洗涤4次去除未结合的FITC,然后把细菌悬浮于PBS中,并调整细菌的终浓度为1×108CFU/mL,每组设6个重复,每个培养孔为1个重复。
1.3.3 Caco-2 细胞的培养
Caco-2细胞用培养瓶培养,在瓶中加入含10%热灭活的胎牛血清和双抗(100 U/mL青霉素和100 μg/mL链霉素)的DMEM 细胞培养液,置于37℃、5%CO2的培养箱中培养,每2 d换1次培养液。待细胞贴壁生长为单层细胞后(5~7 d),用0.25%胰酶消化传代,并用0.4%的台盼蓝染色液染色,用血细胞计数器在显微镜下检测细胞的数量和活性,保证细胞活性在95%以上。
1.3.4 黏附试验
以不加菌的细胞为空白对照,试验组加入1 mL培养液及1 mL荧光标记的乳酸菌、大肠杆菌K88和鼠伤寒沙门氏菌(1×108CFU/mL),将培养板置于37℃,5%CO2的培养箱中分别孵育1 h后,用无菌PBS洗涤3次,将未黏附的菌洗脱。之后每个培养孔中加入0.1 mL胰酶作用5 min,待细胞完全脱落,加入0.4 mL完全培养液终止反应。收集液体,用酶标仪测定其荧光强度,吸收光波长为485 nm,发射光波长为530 nm,测定相对荧光强度,每个组设6个重复,每个培养孔为1个重复。
计算公式:
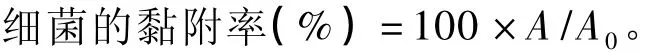
式中:A为乳酸菌、大肠杆菌K88或鼠伤寒沙门氏菌黏附Caco-2细胞并洗脱后细胞悬液的相对荧光强度;A0为乳酸菌、大肠杆菌K88或鼠伤寒沙门氏菌黏附Caco-2细胞前细胞悬液的相对荧光强度。
1.3.5 黏附抑制试验
在96孔板中,每孔加入以上调整好浓度的细胞悬液100 μL。试验设3个处理,通过荧光标记法,采用竞争、排斥和置换试验来研究乳酸菌对病原菌的黏附抑制作用。
1.3.5.1 竞争试验
Caco-2细胞 +乳酸菌(未标记)+致病菌(FITC标记)→5%CO2、37℃共孵育1 h→离心去上清→PBS洗涤3次→PBS重悬→转移至黑色的96孔板,避光静置。
1.3.5.2 排斥试验
Caco-2细胞 +乳酸菌(未标记)→5%CO2、37℃共孵育1 h→离心去上清→加入致病菌(FITC标记)→5%CO2、37℃共孵育1 h→其余操作同1.3.5.1。
1.3.5.3 置换试验
Caco-2细胞 +致病菌(FITC标记)→5%CO2、37℃共孵育1 h→离心去上清→加入乳酸菌(未标记)→5%CO2、37℃共孵育1 h→其余操作同1.3.5.1。
以上处理结束后,各处理的每个培养孔中加入0.1 mL胰酶作用5 min,待细胞完全脱落,加入0.4 mL完全培养液终止反应。收集液体,用酶标仪测定其荧光强度,每组设6个重复,每个培养孔为1个重复。
计算公式:
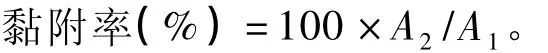
式中:A1为无乳酸菌存在的情况下,黏附在Caco-2细胞上的大肠杆菌K88或鼠伤寒沙门氏菌的相对荧光强度;A2为乳酸菌存在的情况下,黏附在Caco-2细胞上的大肠杆菌K88或鼠伤寒沙门氏菌的相对荧光强度。
1.4 数据分析
试验数据以平均值±标准差表示,采用SPSS 16.0软件作统计学分析,用t检验法进行差异显著性分析,以P<0.05为差异显著,P<0.01为差异极显著。
2 结果与分析
2.1 乳酸菌对Caco-2细胞的黏附作用
由图1可见,参试的8株菌株的黏附率乳杆菌(L2、L3、L1) > 肠球菌(L5、L4) > 致病菌(K88、S)>乳球菌(L6),其中乳酸菌 L1、L2、L3和 L5的黏附率超过50%。可见,研究的这几株益生乳酸菌的黏附性能较好,同时说明乳酸菌对肠上皮细胞的黏附有菌属特异性。分离于健康鸡肠道的乳杆菌L3和来源于人肠道的乳杆菌L1、L2一样有较高的黏附率,这说明乳酸菌对肠上皮细胞的黏附可能无宿主特异性。
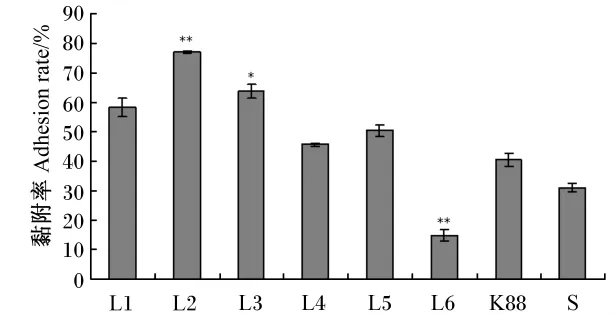
图1 益生乳酸菌对Caco-2细胞的黏附率Fig.1 The adhesion rate of lactic acid bacteria strains to Caco-2 cells
2.2 益生乳酸菌对大肠杆菌K88黏附Caco-2细胞的影响
由表1可知,乳酸菌L1、L2、L3和 L4对大肠杆菌K88黏附Caco-2细胞的影响相似,都能通过置换作用阻止大肠杆菌K88的黏附,相对置换率分别为:42.51%、21.07%、52.49%和37.85%,其中乳酸菌L1、L3和L4的大肠杆菌K88黏附率均极显著降低(P<0.01)。相反,竞争试验或者排斥试验总体上能促进大肠杆菌K88对肠上皮细胞的黏附作用。乳酸菌L5在竞争、排斥和置换试验中都能促进大肠杆菌K88的黏附,乳酸菌L6则能够通过竞争和置换作用降低大肠杆菌K88的黏附,相对降低率分别为6.43%和47.86%。可见,乳酸菌对大肠杆菌K88的黏附影响没有菌属特异性,而且与乳酸菌自身的黏附力也没有必然联系。
2.3 益生乳酸菌对鼠伤寒沙门氏菌黏附Caco-2细胞的影响
由表2可知,所有的乳酸菌都能够通过置换作用减少鼠伤寒沙门氏菌的黏附,乳酸菌L1~L6相对置换率分别为27.38%、11.59%、50.53%、23.16%、57.91%和36.84%,其中 L1、L3~L6的鼠伤寒沙门氏菌黏附率极显著降低(P<0.01)。除了L3外,其他5株益生乳酸菌都通过竞争作用显著(P<0.05)或极显著地(P<0.01)促进其黏附,且所有的乳酸菌都通过排斥作用极显著促进其对Caco-2细胞的黏附(P<0.01)。这表明置换作用是这6株乳酸菌阻止鼠伤寒沙门氏菌黏附Caco-2细胞的主要方式。它们对鼠伤寒沙门氏菌的黏附抑制作用没有菌属特异性,与乳酸菌自身的黏附力也没有关联。
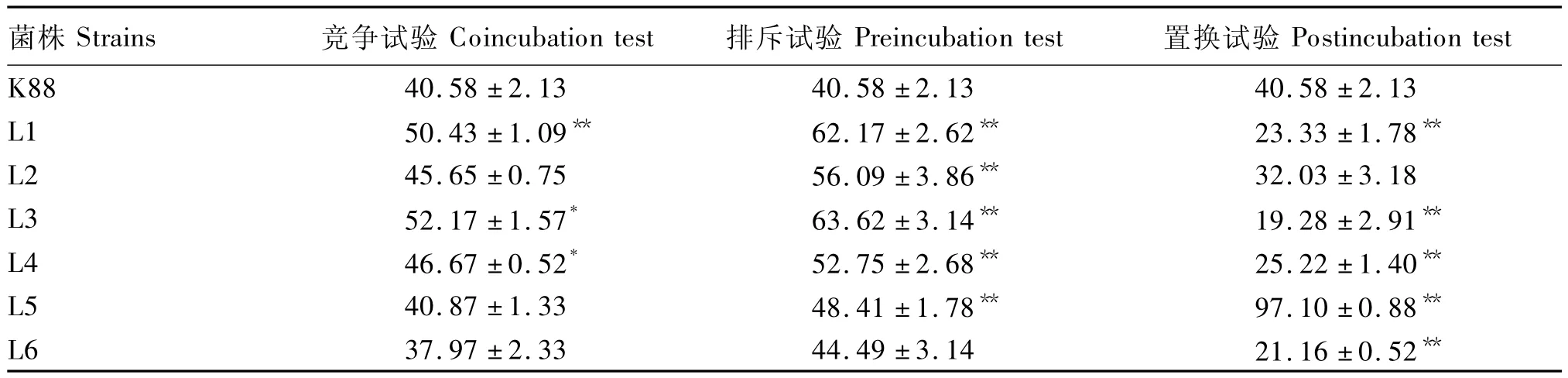
表1 乳酸菌对大肠杆菌K88黏附Caco-2细胞的影响Table 1 Effects of lactic acid bacteria strains on adhesion ability of Escherichia coli K88 to Caco-2 cells %
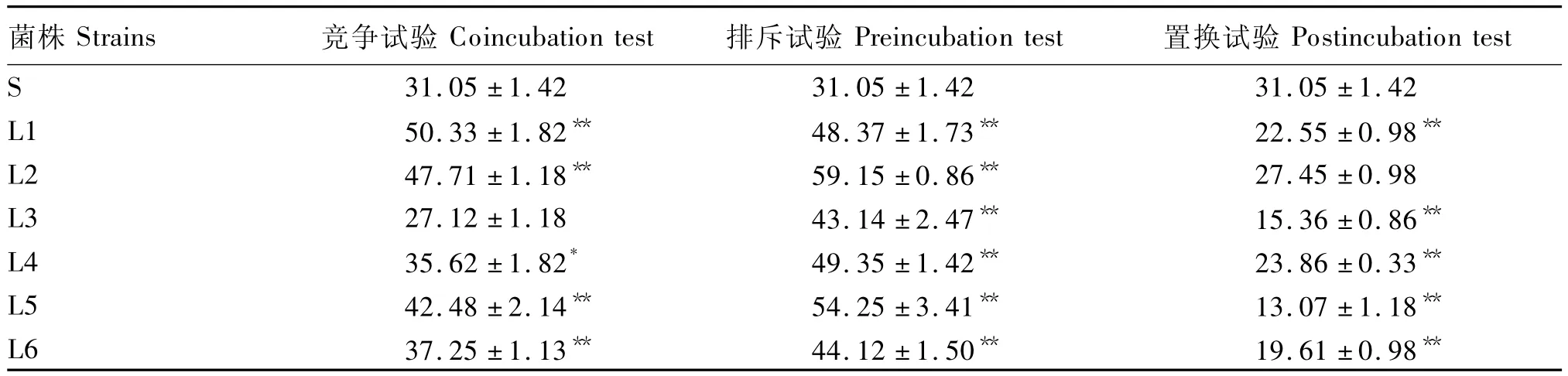
表2 乳酸菌对鼠伤寒沙门氏菌黏附Caco-2细胞的影响Table 2 Effects of lactic acid bacteria strains on adhesion ability of S.typhimurium to Caco-2 cells %
3 讨论
随着抗生素在全球动物生产中的禁用,益生菌成为研究的热点,乳酸菌作为益生菌的一种,因其能够改善胃肠道功能、提高动物免疫力和生产性能等,在动物生产中得到了极大的推广和应用。对于益生菌的作用机理,目前普遍认为黏附是其发挥益生作用的前提,也是细菌与宿主相互作用的第1步。菌群在肠道内的定植是通过细菌的黏附作用来实现的,乳酸菌进入肠道能否黏附于宿主某段肠道的黏膜细胞表面,形成稳定的优势菌群,是决定这种益生素实际效果的重要因素。Gopal等[8]报道3株饲用乳酸菌菌株对人肠道上皮细胞系 HT-29、Caco-2 和 HT29-MTX 细胞有极强的黏附力,而且3株乳酸菌都能降低大肠杆菌O157∶H7对肠细胞的侵袭能力和细胞结合能力。陈臣等[9]研究发现3株乳杆菌对Caco-2细胞均有较好的黏附能力,尤其是植物乳杆菌ST-Ⅲ,其黏附率高于被深入研究的鼠李糖乳杆菌。本试验发现,试验菌株黏附率从高到低依次为乳杆菌>肠球菌>致病菌>乳球菌,说明乳酸菌在体外对肠上皮细胞有较好的黏附性能,与上述报道基本一致。同时说明乳酸菌对Caco-2细胞的黏附有菌属特异性,这与Nissen等[10]报道的结果一致。这可能与黏附素有很大的关系,黏附素是一种与黏附相关的分泌型蛋白质,它可介导菌体与细胞膜受体的连接,此类分泌型蛋白质分子的水平在很大程度上可能不尽一致,由此表现出乳酸菌黏附性能的差异。本试验发现分离于健康鸡肠道的乳杆菌L3和来源于人肠道的乳杆菌L1、L2对Caco-2细胞的黏附率没有显著差异。王斌等[11]认为,乳杆菌对肠上皮细胞的黏附有宿主特异性,而与此相反,李平兰等[12]却发现,分离自人粪便与猪粪便的同属乳酸菌黏附HT-29细胞的能力没有显著差异。本试验结果提示乳酸菌的黏附可能无宿主特异性,与李平兰等[12]的研究结果一致,但要确定乳酸菌是否具有宿主特异性还需进一步的试验验证。本试验发现几乎所有乳酸菌均能通过置换作用抑制大肠杆菌K88和鼠伤寒沙门氏菌对肠上皮细胞的黏附,但是通过竞争作用和排斥作用不但不能抑制反而促进病原菌的黏附。虽然不同种属和菌株的益生乳酸菌对肠道致病菌的黏附抑制作用不同[13],但是很多益生乳酸菌抑制肠致病菌黏附肠上皮细胞都是通过排斥、竞争或置换作用实现。Matijašic'等[14]研究发现,格氏乳杆菌通过排斥和竞争作用抑制大肠杆菌K88黏附Caco-2细胞。郭彤等[15]报道,嗜酸乳杆菌能够通过排斥、竞争或置换作用明显抑制大肠杆菌K88和猪霍乱沙门氏菌黏附 Caco-2 细胞。Tuomola 等[16]和 Collado等[17]分别报道过乳酸杆菌和双歧杆菌对病原菌黏附的促进作用,但具体原因尚不清楚。但是体外细胞试验是在人为的简单的环境中进行,与体内真实复杂的环境还是有区别的。在宿主的肠道内,肠上皮细胞表面还覆盖有一层由一些糖蛋白类物质组成的黏液层,黏蛋白是其中主要的一种糖蛋白,其上分布有肠菌群的一些特异性结合位点。益生菌的黏附特点可能与其细胞壁成分或代谢产物有关。益生细菌细胞壁的成分(如脂膜酸)及其含量或者代谢物质及其分泌量,以及细菌表面的特性都影响其黏附力[18-19],从而导致其对病原菌黏附性能的影响不同。Mangell等[20]认为有黏附力的益生菌不容易因肠蠕动而随食糜排出体外,从而容易在肠道中定植,构成抵抗致病菌入侵的微生物屏障,发挥更持久的保护作用。因此,黏附力一直是受人关注的肠道益生菌特性之一。本试验中的乳酸菌菌株黏附性能较好,但是关于其对病原菌的黏附抑制作用及其乳酸菌的黏附是否具有宿主特异性还有待进一步研究以得到更加明确的结果。对于乳酸菌的黏附机理也还需要进一步的研究,以期能在动物生产上更好地利用黏附特性来挑选乳酸菌进行生产试验,从而对其提高动物机体免疫力的能力有更好的评价。
4 结论
①乳酸菌对肠上皮细胞有较高的黏附率,尤其是乳杆菌。
②乳酸菌对肠上皮细胞的黏附有菌属特异性。
③乳酸菌对大肠杆菌K88和鼠伤寒沙门氏菌的黏附抑制作用主要通过置换方式,但没有菌属特异性,且与自身的黏附力也没有必然的联系。
[1]COTTER P A,MILLER J F.Triggering bacterial virulence[J].Science,1996,273:1183 - 1185.
[2]COLLINS J K,THORNTON G,SULLIVAN G O.Selection of probiotic strains for human applications[J].International Dairy Journal,1998,8:487 - 490.
[3]范小兵,周丛,杭晓敏,等.昂立植物乳杆菌黏附肠上皮细胞的研究[J].中国微生态学杂志,2004,16(6):353-355.
[4]王斌,李秋荣,刘放南,等.1株肠道高黏附性乳杆菌的分离鉴定[J].中国生物制品学杂志,2008,21(6):473-477.
[5]CANDELA M,PERNA F,CARNEVALI P,et al.Interaction of probiotic Lactobacillus and Bifidobacterium strains with human intestinal epithelial cells:adhesion properties,competition against enteropathogens and modulation of IL-8 production[J].International Journal of Food Microbiology,2008,125(3):286 -292.
[6]VINDEROLA C G,MEDICI M,PERDIGÓN G.Relationship between interaction sites in the gut,hydrophobicity,mucosal immunomodulating capacities and cell wall protein profiles in indigenous and exogenous bacteria[J].Journal of Applied Microbiology,2004,96(2):230-243.
[7]BONAR A,CHMIELA M,RUDNICKA W,et al.Mannose-binding lectin enhances the attachment and phagocytosis of mycobacteria in vitro[J].Archivum Immunologiae et Therapia Experimentalis,2005,53:437-441.
[8]GOPAL P K,PRASAD J,SMART J,et al.In vitro adherence properties of Lactobacillus rhamnosus DR20 and Bifidobacterium lactis DR10 strains and their antagonistic activity against an enterotoxigenic Escherichia coli[J].International Journal of Food Microbiology,2001,67(3):207 -216.
[9]陈臣,郭本恒,陈卫,等.三株益生菌黏附性质及机制的初步研究[J].中国微生态学杂志,2007,19(6):492-494.
[10]NISSEN L,CHINGWARU W,SGORBATI B,et al.Gut health promoting activity of new putative probiotic/protective Lactobacillus spp.strains:a functional study in the small intestinal cell model[J].International Journal of Food Microbiology,2009,135(3):288-294.
[11]王斌.乳杆菌黏附机制的研究进展[J].国际检验医学杂志,2006,27(3):224 -226.
[12]李平兰,杨华,张篪.乳酸菌体外黏附人结肠腺癌细胞系HT-29细胞的研究[J].中国农业大学学报,2002,7(1):19 -22.
[13]GUEIMONDE M,JALONEN L,HE F,et al.Adhesion and competitive inhibition and displacement of human enteropathogens by selected Lactobacilli[J].Food Research International,2006,39(4):467 - 471.
[14]MATIJAŠIC'B B,NARAT M,PETERNEL M Z,et al.Ability of Lactobacillus gasseri K7 to inhibit Escherichia coli adhesion in vitro on Caco-2 cells and ex vivo on pigs’jejunal tissue[J].International Journal of Food Microbiology,2006,107(1):92 -96.
[15]郭彤,胥保华,马玉龙,等.嗜酸乳杆菌和两歧双歧杆菌体外黏附Caco-2细胞及其对病原菌黏附性能的影响[J].中国兽医学报,2008,28(5):527 -531.
[16]TUOMOLA E M,OUWEHAND A C,SALMINEN S J.The effect of probiotic bacteria on the adhesion of pathogens to human intestinal mucus[J].FEMS Immunology & Medical Microbiology,1999,26(2):137-142.
[17]COLLADO M C,GUEIMONDE M,HERNÁNDEZ M,et al.Adhesion of selected Bifidobacterium strains to human intestinal mucus and the role of adhesion in enteropathogen exclusion[J].Journal of Food Protection,2005,68(12):2672 -2678.
[18]陈臣,郭本恒,王荫榆,等.植物乳杆菌ST-Ⅲ黏附肠上皮样细胞系Caco-2性质的研究[J].乳业科学与技术,2008,31(2):51 -55.
[19]GRANATO D,BERGONZELLI G E,PRIDMORE R D,et al.Cell surface-associated elongation factor Tu mediates the attachment of Lactobacillus johnsonii NCC533(La1)to human intestinal cells and mucins[J].Infection and Immunity,2004,72(4):2160 -2169.
[20]MANGELL P,LENNERNAS P,WANG M,et al.Adhesive capability of Lactobacillus plantarum 299v is important for preventing bacterial translocation in endotoxemic rats[J].Acta Pathologica,Microbiologica et Immunologica Scandinavica,2006,114(9):611 -618.
猜你喜欢
石油沥青(2023年4期)2023-09-21 07:18:12
石油沥青(2023年4期)2023-02-27 12:42:02
科学(2020年3期)2020-11-26 08:18:22
当代水产(2020年3期)2020-06-15 12:03:02
上海公路(2019年1期)2019-06-18 11:05:06
食品与机械(2019年1期)2019-03-30 01:14:36
兽医导刊(2016年12期)2016-05-17 03:51:45
实用皮肤病学杂志(2015年4期)2015-12-22 11:21:42
医学研究杂志(2015年12期)2015-06-10 06:57:46
特产研究(2014年4期)2014-04-10 12:54:12
