
Ⅰ类整合子在耐多药产ESBLs大肠埃希菌中的分布探讨*
2012-08-13 09:45李佩波蔡敏泓黄永茂邹永胜向成玉
重庆医学 2012年6期
李佩波,蔡敏泓,黄永茂,林 雁,邹永胜,钟 利,陈 枫,陈 庄,向成玉
(泸州医学院附属医院感染科,四川 泸州 646000)
大肠埃希菌属于肠杆菌科,为革兰阴性短杆菌,属于条件致病菌,是临床上常见的细菌感染之一。近年来,随着β-内酰胺类、氨基糖苷类、喹诺酮类等抗菌药物在临床上广泛应用、不合理使用以及滥用,导致细菌出现了不同程度的耐药,甚至出现了交叉耐药和多重耐药的特点[1]。Ⅰ类整合子在大肠埃希菌中广泛存在,整合子相关耐药基因在该菌耐药性的形成和播散中发挥重要作用[2]。近年来研究发现大肠埃希菌耐药情况严重,尤其是产ESBLs酶菌株更突出[3]。本研究对临床分离的71株大肠埃希菌进行耐药分析、ESBLs鉴定和Ⅰ类整合子遗传标记检测,并探讨其相关性,现报道如下。
1 材料与方法
1.1 材料 实验菌株:71株大肠埃希菌临床分离株(不含同一病例相同部位重复分离株)来自泸州医学院附属医院2007年10月至2008年10月期间感染大肠埃希菌的患者。其中呼吸道感染29例,泌尿系统感染18例,外科切口感染14例,全身感染8例,其他2例;全部菌株均重新经VITEK系统(BioMerieux,法国)鉴定确认。标准菌株:大肠埃希菌 ATCC 25922,购自国家临床检验中心。
1.2 方法
1.2.1 药物敏感试验 采用美国全国临床检验标准委员会(NCCLS)推荐的纸片扩散法(K-B法)进行临床常用3类6种(LVE、CIP、FOX、S、A)抗菌药物的敏感性检测,并根据 NCCLS 2006要求进行敏感性判断。
1.2.2 ESBLs鉴定 初步筛选试验:选用头孢噻肟、头孢他啶、氨曲南、头孢曲松药敏纸片(杭州天和微生物试剂公司)对71株大肠埃希菌进行产ESBLs筛选试验,凡头孢他啶的抑菌环直径小于或等于22mm,头孢曲松小于或等于25mm,氨曲南或头孢噻肟小于或等于27mm,则提示为可疑产ESBLs菌株。纸片确认试验:将初筛可疑产ESBLs菌株同时用头孢他啶和头孢他啶加克拉维酸、头孢噻肟和头孢噻肟加克拉维酸作药敏试验,对任何一组药物,加克拉维酸前后抑菌环直径相差大于或等于5mm可确认为产ESBLs菌株。
1.2.3 PCR技术检测基因 采用Biospin细菌基因组DNA试剂盒(上海生工生物技术公司)提取大肠埃希菌DNA。根据Ⅰ类整合子遗传标记qacE△sul1设计引物(上海生工生物技术公司提供)。上游引物基因序列P1:TAG CGA GGG CTT TAC TAA GC;下游引物基因序列P2:ATT CAG AAT GCC GAA CAC CG。
以提取的大肠埃希菌DNA为反应模板进行PCR扩增。所有PCR反应体系均为25μL,其中2×Taq PCR MasterMix组成包括:0.1U/μL聚合酶,500mmol/L dNTP,20mmol/L Tris-HCl(pH 8.3),100mmol/L KCl,3mmol/L MgCl2等。PCR扩增条件:94℃预变性3min,94℃变性30s,55℃退火30s,72℃延伸60s,共30个循环,然后72℃延伸5min。将扩增产物进行琼脂糖凝胶电泳、紫外线检测仪观察结果,并将扩增产物送至上海生工生物技术公司进行基因测序。
1.3 统计学处理 所有实验结果均采用SPSS13.0录入建立数据库并行统计分析,统计方法采用χ2检验。当T<1或N<40时用确切概率法计算P值;若1≤T<5且N≥40时用连续校正法处理。
2 结 果
2.1 药物敏感试验结果 71株临床分离的大肠埃希菌对6种抗生素耐药情况分别为:左氧氟沙星45株、环丙沙星51株、头孢噻肟44株、头孢西丁9株、链霉素41株、阿米卡星6株。其中耐药率最高为环丙沙星72.86%,最低为阿米卡星8.57%。不同耐药模式中以耐多药最多为45株(64.29%),全敏最少为7株(9.86%)。
2.2 ESBLs鉴定结果 产ESBLs酶表型筛选试验初步筛出55株(77.46%),可疑产ESBLs菌株。通过进一步确认试验在58株疑产ESBLs菌株中共检出产ESBLs菌35株(49.30%)。
2.3 大肠埃希菌中整合子遗传标记基因的PCR法检测情况
2.3.1 整合子遗传标记在不同耐药模式中检出结果见表1。

表1 各种耐药模式中整合子遗传标记基因阳性率比较
2.3.2 整合子遗传标志在产ESBLs酶菌株中检出结果 见表2。

表2 产ESBLs酶大肠埃希菌不同耐药模式中整合子遗传标记检出情况
2.3.3 整合子遗传标记在非产ESBLs酶大肠埃希菌中检出结果 见表3。

表3 非产ESBLs酶菌株不同耐药模式中整合子遗传标记基因阳性率的比较
2.4 整合子遗传标记qacE△sul1电泳图片和基因测序结果
将本实验中Ⅰ类整合子遗传标记(qacE△sul1)PCR扩增产物序列测序后的基因在BLAST上进行比对发现与基因库中已登陆的基因相同,整合子遗传标记qacE△sul1电泳图见图1。
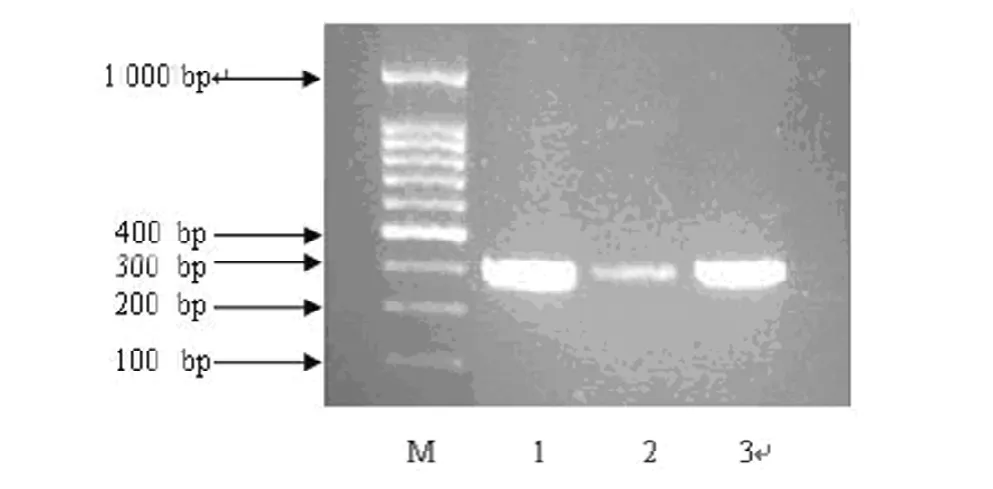
图1 整合子遗传标记qacE△sul1电泳图
3 讨 论
本研究中菌株对左氧氟沙星、环丙沙星、头孢噻肟、链霉素耐药情况严重,尤其是在多耐药模式中更为突出,以CTX+LVE/CIP+S模式常见。但对头孢西丁和阿米卡星仍有较高的敏感性,这与头孢西丁对β-内酰胺酶高度稳定,阿米卡星对氨基糖苷类钝化酶稳定的抗菌作用特点有关。
大肠埃希菌产ESBLs在不同国家的检出有明显差异,如土耳其大肠埃希菌的ESBLs发生率仅为12%[3];而国内近5年大肠埃希菌产ESBLs菌株检出率从46.0% 上升至63.4%,呈逐年增高的趋势[5]。CTX-M(以高度水解头孢噻肟为特征已dn为国内外流行最广的ESBLs[6])。本研究中大肠埃希菌ESBLs检出率为49.30%(36株),与文献所报道基本一致。
基因盒-整合子系统多见于革兰阴性杆菌,以肠杆菌、铜绿假单胞菌为主,是细菌尤其是革兰阴性菌多重耐药快速发展的重要原因[7]。依据整合酶基因序列的不同,将整合子分为6类,亦有文献报道为8类[8],目前报道较多的仅有4类。Ⅰ~Ⅲ类整合子已被证明与细菌耐药性有关,为耐药整合子,其中第Ⅰ、Ⅱ类整合子是最常见捕获和表达耐药基因的整合子[9]。近年来,国外大量文献报道整合子可介导病原体各种药基因获得整合子病原菌可表现为多药耐药[10-13]。QacE△1-sul1是Ⅰ类整合子的遗传标记,实质是qacE△1基因和sul1基因组成的重叠基因,位于Ⅰ类整合子3′保守区。Dahmen等[14]研究提示:6种不同肠杆菌中I类整合子基因具有较高的检出率,整合子遗传标记以sul1常见,sul1和sul2的重叠显现明显。有学者对血培养中的大肠埃希菌进行研究发现,几乎有一半菌株中整合子及Sul基因阳性,其中以经典类整合子qacE△1-sul1最多。
本研究结果提示,Ⅰ类整合子在大肠埃希菌中有高的检出率(78.57%),与文献报道相近。实验结果提示:大肠埃希菌产ESBLs酶且整合子阳性者,出现耐药概率比单纯携带整合子或者产ESBLs酶的菌株要大,尤其在多耐药模式中更为明显。
[1]李佩波,黄永茂.大肠埃细菌耐药及机制研究进展[J].西南军医,2011,13(1):95-97.
[2]Diaz-Mejta JJ,Amabile-Cuevas CF,Rosas I,et al.An analysis of the evolutionary relationships of integron integrases,with emphasis on the prevalence of class 1integrons in Escherichia coli isolates from clinical and environmental origins[J].Microbiology,2008,154(1):94-102.
[3]褚云卓,田素飞,王玉珏,等.产超广谱β-内酰胺酶大肠埃希菌的临床调查中[J].国实验诊断学,2009,13(10):1423-1425.
[4]Qzqunes I,Erben N,Kiremitci A,et al.The prevalence of extended-spectrum beta-latamase-producing Escherchia coli and Klebsiella pneumoniae in clinical isolates and risk factors[J].Scan Med J,2006,27(5):608-612.
[5]战榕,陈 菁,黄心宏,等.2004~2009年我院常见病原菌的耐药性变迁分析[J].福建医科大学学报,2010,44(3),205-209.
[6]Livermore DM,Hawkey PM.CTX-M:changing the faue of ESBls in the UK[J].J Amimicrob Chemother,2005,56(3):451-454.
[7]吴志鹃,黄永茂.基因盒-整合子系统介导细菌多重耐药的研究进展[J].国外医药抗生素分册,2010,31(1):34-37.
[8]Barlow RS,Pemberton JM,Desmarchelier PM,et al.Isolation and characterization of integron-containing bacteria without antibiotic selection.[J]Antimicrob Agents Che-mother,2004,48(3):28-38.
[9]Weldhagen GF.Integrons andβ-lactamases-a novel perspective on resistance[J].Int J Antimicrob Agents,2004,23(6):556-562.
[10]林宁,孙海平.多药耐药大肠埃细菌整合子、转座子遗传标记研究[J].中华医院感染学杂志,2008,18(10):1361-1363.
[11]Kang SG,Lee DY,Mshin SJ,et al.Changes in patterns of an timicorobiol susceptibility and class I grom carriage anong Escheriehia coli isolates[J].J Vet Sec,2005,6(3):201-205.
[12]Doi Y,Adams JM,Yamane K,et al.Identification of 16s RNA methlase-producing acinetobacter baumannii clinican strains in North America[J].Antimicrob Agents Chemother,2007,51(11):4209-4210.
[13]糜祖煌,秦玲.多耐药鲍氏不动杆菌5类抗菌药物机制研究[J].中华医院感染学杂志,2008,18(7):901-904.
[14]Dahmen S,Mansour W,Boujuafar N,et al.Distribution of cotrimoxazole resistance genes associated with class 1integrons in clinical isolates of enterobacteriaceae in a university hospital in tunisia[J].Microb Drug Resist,2010,16(1):43-47.
猜你喜欢
云南医药(2021年3期)2021-07-21
爱你·健康读本(2020年8期)2020-08-23
爱你(2020年22期)2020-08-18
中国医学装备(2016年6期)2016-12-01
中华灾害救援医学(2015年7期)2016-01-07
现代检验医学杂志(2015年2期)2015-02-06
中国药业(2014年12期)2014-06-06
中国合理用药探索(2014年11期)2014-03-11
当代畜禽养殖业(2014年4期)2014-02-27
现代检验医学杂志(2014年1期)2014-02-06
