p53异构体和MDM2、PTEN在胃癌组织中表达的相关性
2012-08-05 06:03季万胜刘俊衡刘丹丹高志星
山东医药 2012年13期
孙 娜,季万胜,刘俊衡,刘丹丹,高志星
(潍坊医学院,山东潍坊261041)
胃癌是指起源于胃黏膜上皮的恶性肿瘤,目前认为胃癌的发生发展是多环节、多因素、多基因相互作用的结果。p53抑癌基因功能的失活在胃癌的发生发展过程中起重要作用。但最近的统计数据表明,中国内地胃癌 p53的突变率为30.6%[1],提示部分胃癌的发生并没有伴随着p53的高突变率。研究显示,p53的生物化学和生物学功能是可以通过p53异构体和全长p53蛋白之间的相互作用来调节的。因此,研究p53异构体在胃癌组织中的表达模式、异构体对野生型p53功能的影响、特别是对p53下游分子表达水平的影响,有可能发现胃癌发生的新机制。我们采用PCR、免疫组化方法检测了胃癌组织和癌旁胃组织中 p53异构体、双微基因2(MDM2)、同源性磷酸酶张力蛋白(PTEN)的表达情况,并探讨其在胃癌发生、发展中的作用及它们之间的相互关系,为胃癌的早期和临床诊断提供更多的新思路和新方法。
1 资料与方法
1.1 临床资料 收集2009年11月~2011年2月于潍坊医学院附属医院普外科行胃癌根治术的30例胃癌患者的手术切除标本[包括胃癌组织和癌旁胃组织(距离正常胃癌组织5 cm)]。患者在术前均未接受任何放化疗。男21例、女9例,年龄40~82岁、平均56岁。所取肿瘤组织均经术后病理诊断为胃癌,所取癌旁胃组织均经病理诊断未检测到癌细胞。
1.2 方法
1.2.1 PCR法检测胃癌组织和癌旁胃组织中p53β、p53γ、Δ133p53、Δ133p53β、Δ133p53γ 的表达情况 应用UNIQ-10柱式Trizol总RNA提取试剂盒提取胃癌组织和癌旁胃组织的总RNA。胃组织RNA逆转录反应:按照MMLV第一链cDNA合成试剂盒说明书及相关文献操作,以试剂盒内的Oligo-p(dT)18Primer为随机引物,以M-MuLV RT酶为逆转录酶进行体外逆转录反应。胃组织中 p53β、p53γ、Δ133p53、Δ133p53β、Δ133p53γ 五种 p53 异构体PCR扩增。总RNA提取试剂盒、cDNA合成试剂盒、PCR扩增试剂盒、引物购自Sangon上海生工,BIO-RAD MyCyder PCR仪购自美国伯乐公司。PCR扩增产物进行琼脂糖凝胶电泳后,将凝胶放入购自美国UVP的BioSpectrum AC凝胶成像系统扫描并拍照,并对电泳结果进行判定。
1.2.2 免疫组化PV-9000两步法检测胃癌、癌旁组织中MDM2和PTEN的表达情况 标本经10%甲醛固定,石蜡包埋,常规HE染色,病理确诊,免疫组化步骤按说明书进行。鼠抗人MDM2和PTEN单克隆抗体均购自北京中杉金桥,均为3 mL工作液。以PBS代替一抗作阴性对照。每张切片随机选择5个视野,共计数100个肿瘤细胞,分别计算2种抗体的阳性细胞百分率。由2位以上病理医生对免疫组织化学染色进行评价,依据镜下切片中阳性染色细胞占瘤细胞总数的比例分为2级,-:无染色或染色阳性细胞数≤10%,+:染色阳性细胞数>10%。
1.2.3 统计学方法 采用SPSS16.0统计软件,数据以百分比表示,比较行χ2检验,应用Spearman’s等级相关进行相关性分析。P≤0.05为差异有统计学意义。
2 结果
2.1 p53异构体在胃癌组织和癌旁胃组织中的表达 在30例胃癌组织和癌旁胃组织中均未检测到p53γ、Δ133p53β、Δ133p53γ 这三种异构体的 mRNA表达。Δ133p53的表达结果:在30例胃癌组织中有21例表达,占70.0%(21/30);在30例癌旁胃组织中有7例表达,占 23.3%(7/30);胃癌组织中Δ133p53表达高于癌旁胃组织(P <0.01)。p53β的表达结果:在30例胃癌组织中有8例表达,占26.7%(8/30);在30例癌旁胃组织中有21例表达,占70.0%(21/30),胃癌组织p53β的表达低于癌旁胃组织(P <0.01)。
2.2 MDM2、PTEN在胃癌组织和癌旁胃组织中的表达 MDM2的表达结果:在30例胃癌组织中有19例表达,占63.3%(19/30);在30例癌旁胃组织中有7例表达,占23.3%(7/30),MDM2在胃癌组织中表达高于癌旁胃组织(P<0.01)。PTEN的表达结果:在30例胃癌组织中有16例表达,占53.3%(16/30);在30例癌旁胃组织中有26例表达,占86.7%(26/30),胃癌组织PTEN的表达低于癌旁胃组织(P <0.01)。
2.3 p53异构体和MDM2、PTEN表达的相关性分析 Spearman’s相关性分析显示,Δ133p53和MDM2,p53β和PTEN在胃癌中表达呈正相关关系(r=0.408,P=0.025;r=0.413,P=0.023);Δ133p53和PTEN,p53β和MDM2在胃癌中表达呈负相关关系(r= -0.467,P=0.009;r= -0.480,P=0.007)。见表 1、2。
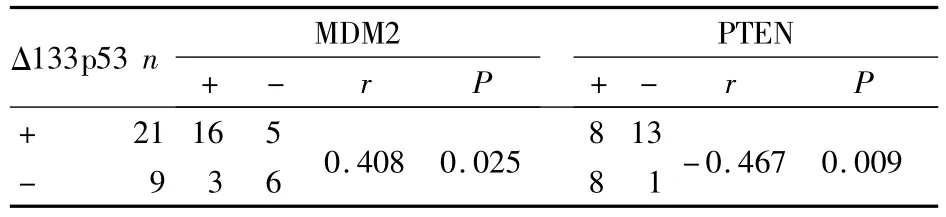
表1 胃癌组织中Δ133p53与MDM2、PTEN相关性分析

表2 胃癌组织中p53β与MDM2、PTEN相关性分析
3 讨论
抑癌基因p53的功能失活在胃癌的发生发展过程中起重要作用,p53异构体的发现为胃癌发生机制的研究提供了新方向。p53异构体的活性主要通过改变内部启动子的活性、p53 mRNA的剪接及p53异构体的亚细胞定位来实现[2]。最近研究发现,人类p53异构体(Δ133p53和p53β)能够组成一内源性的调节机制,并且能够作用于p53介导的复制细胞的衰老。在人类正常成纤维细胞中,p53β的诱导生成和Δ133p53的减少与复制细胞的衰老有关,这种衰老与癌基因所致的衰老不同。在结肠腺癌中可以观察到Δ133p53表达水平升高和p53β表达水平下降,这可能是在从腺瘤到癌的进展过程中,逃脱了衰老障碍的一种信号[3],即天然存在的p53异构体为复制细胞的衰老进行生理学上的调节。随着体内试验证据表明,p53异构体特异性表达的建立和缺失能够分别诱导和逃脱衰老状态,如果打破这种衰老机制则可以诱导肿瘤的生成。本研究显示,在胃癌组织中发现Δ133p53 mRNA水平增高,p53βmRNA水平降低,与文献报道一致。
p53基因是人类发现最早的抑癌基因,MDM2高表达呈现癌基因的功能,其表达产物在细胞核内可以与野生型和突变型p53结合使其功能失活。p53和MDM2之间存在着自我反馈环,该反馈环使细胞内的MDM2/p53比率保持恒定[4]。通过研究p53-MDM2自我反馈环,发现MDM2与p53结合后可使p53泛素化,泛素化的p53被蛋白酶体降解而直接被清除[5,6]。正确的亚细胞定位对于p53的活性十分重要,p53被激活后进人核内,激活其靶基因的转录,之后回到胞质中被降解。最近发现,p53向核外转运可能以MDM2依赖性的方式进行控制,因为MDM2中含有的NLS和NES保证了MDM2可以穿梭于胞核与胞质之间,核内MDM2将p53捕获,通过MDM2上的NLS将两种蛋白质运出细胞核。如果细胞将MDM2修饰或改变核输出装置使MDM2滞留在核,p53将不被降解。PTEN是具有磷酸酶活性的抑癌基因,该基因的编码产物能够使被PI3K激活的细胞内第二信使PIP3去磷酸化,从而影响细胞内 PI3K/Akt信号转导途径[7,8]。MDM2 是一原癌基因,最近研究表明,PI3K/Akt信号转导途径可以激活细胞的磷酸化过程,从而导致MDM2癌蛋白由胞质进入胞核,而PTEN可以通过对该信号转导途径的影响,阻止MDM2癌蛋白由胞质进入胞核,从而加速该蛋白的降解[7,9]。由此PTEN 参与到p53-MDM2自我反馈环中,形成了PTEN-MDM2-p53网络环路,影响着肿瘤的生成。
本研究结果显示,在胃癌组织和癌旁胃组织中p53异构体的表达情况不同,能够检测到Δ133p53、p53β 表达,未检测到 p53γ、Δ133p53β、Δ133p53γ 的表达。Δ133p53、MDM2在胃癌组织中的表达高于癌旁胃组织,p53β、PTEN在癌旁胃组织中的表达高于胃癌组织。相关性分析显示,Δ133p53和MDM2,p53β和 PTEN在胃癌中的表达均呈正相关;Δ133p53和PTEN,p53β和MDM2在胃癌中的表达均呈负相关。由此推测,在胃癌的发生、发展过程中,Δ133p53、p53β通过调节 p53活性,并以 p53为中介,影响了PTEN-MDM2-p53网络环路中PTEN、MDM2蛋白的表达,并提示 p53异构体中的Δ133p53可能是通过降低PTEN、p53的抑癌蛋白表达及提高MDM2促癌蛋白的表达而发挥促癌作用的;而p53β可能是通过提高PTEN、p53的抑癌蛋白表达及降低MDM2促癌蛋白的表达,调控PTENMDM2-p53网络环路从而促进胃癌的发生。
[1]Ghosh A,Stewart D,Matlashewski G.Regulation of human p53 activity and cell localization by altemative splicing[J].Mol Cell Biol,2004,24(18):7987-7997.
[2]Anensen N,Oyan AM,Bourdon JC,et al.Adistinet p53 protein isoform signature reflects the onset of induction chemotherapy for acute myeloid leukemia[J].Clin Cancer Res,2006,12(13):398-392.
[3]Candeias MM,Powell DJ,Roubalova E,et al.Expression of p53 and p53/47 are controlled by alternative mechanisms of messenger RNA translation initiation[J].Oncogene,2006,25(52):6936-6947.
[4] Watanabe T,Ichikawa A,Saito H,et al.Overexpression of the MDM2 oncogene in leukemia and lymphoma[J].Leuk Lymphoma,1996,21(5-6):391-397.
[5]Freedman DA,Wu L,Levine AJ.Functions of the MDM2 oncoprotein[J].Cell Mol Life Sci,1999,55(1):96-107.
[6]Juven-Gershon T,Oren M.MDM2:the ups and downs[J].Mol Med,1999,5(2):71-83.
[7]Yunqing L,Fadila G.PTEN has tumor-promoting properties in the setting of gain-of-function p53 mutations[J].Cancer Res,2008,68(6):1723-1731.
[8]周旭,易继林,刘恩宇,等.PTEN基因在肝细胞肝癌中的表达及其意义[J].肿瘤防治杂志,2005,12(4):286-290.
[9]Choi HJ,Chung TW.Ganglioside GM3 modulates tumor suppressor PTEN-mediated cell cycle progression——transcriptional induction of p21(WAF1)and p27(kip1)by inhibition of PI-3K/AKT pathway[J].Glycobiology,2006,16(7):573-583.
猜你喜欢
系统仿真技术(2022年4期)2023-01-17
云南化工(2021年8期)2021-12-21
天津医药(2016年9期)2016-10-20
国外医药(抗生素分册)(2016年4期)2016-07-12
中国男科学杂志(2016年9期)2016-03-20
信息记录材料(2016年4期)2016-03-11
中国医疗美容(2015年1期)2015-07-12
医学研究杂志(2015年9期)2015-07-01
中国老年学杂志(2015年16期)2015-03-05
癌变·畸变·突变(2015年4期)2015-02-27