
舟山群岛普陀樟天然群落特征研究
2012-07-30 10:01:36高大海贺位忠吴初平
浙江林业科技 2012年6期
陈 斌,高大海,贺位忠,鲁 专,吴初平
(1. 浙江省舟山市林业科学研究院,浙江 定海 316000;2. 浙江省舟山市农林局,浙江 定海 316021;3. 浙江省林业科学研究院,浙江 杭州 310023)
普陀樟(Cinnamonum japonicumvar.chenii)为国家二级重点保护植物,系樟科Lauraceae常绿乔木,具有优良的观赏性状,幼树耐庇荫。由于其根系发达,且耐热、耐寒、耐旱、抗风,生性强健,是庭院及滨海地区园林绿化中不可多得的优良树种。目前国内对普陀樟开展研究较少,主要集中在育苗、引种、天然林资源调查等方面,而未见普陀樟群落结构特征方面的研究报道。近年来在舟山地区作为主要造林树种开始大量应用。
由于普陀樟自然分布特征明显,应用前景广阔,研究其群落特征显得尤为必要。调查组于2011年6-7月进行了外业调查,调查发现除在以前文献报道中所记载的天然群落分布点外[1~2],桃花岛存在普陀樟天然分布,且在之前文献中均未设置标准样地进行群落研究。为了进一步研究普陀樟群落特征,本文通过对普陀樟2类6个群落的研究,阐明普陀樟天然林群落结构特征、群落的物种多样性所表现的物种竞争性,有助于揭示其分布原因,为普陀樟的保护和开发提供依据。
1 自然概况与研究方法
1.1 自然概况
普陀樟主要天然分布与浙江省舟山市普陀区,全区总面积 6728 km2,29° 32′ ~ 30° 28′ N,121° 56′ ~ 123°14′,其中海域面积6269.4 km,陆地面积458.6 km,海岸线总长831.43 km,共有大小岛屿455个,最高峰为桃花岛对峙山,海拔544.5 m。
普陀属北亚热带南缘季风海洋型气候,常年温和湿润,冬暖夏凉,光照充足。年平均气温 16.1℃,极端最高温度38.2℃,极端最低温度-6.5℃,最冷为1月,平均温度5.5℃,8月最热,月平均气温26.9℃,常年日照时数2150.7 h,全年≥10℃平均活动积温5009.6℃,无霜期279 d。年平均降水量1200 mm。年平均相对湿度80%,属湿润或半湿润地区。
1.2 调查方法
1.2.1 样地选择与设置 在查阅相关文献资料后,除对记录岛屿调查外,还对周边岛屿进行了全面踏查,基本掌握了普陀樟的自然分布状况,通过踏查,发现桃花岛与朱家尖还存在2处未记录的普陀樟小片天然分布。在普陀区的朱家尖岛、桃花岛、普陀山岛、络迦山、悬鹁鸪岛5个岛,选择二类群落(一为唯一的3个作为上层木出现的群落,一为下层木存在),设置6个样地进行调查,以分析普陀樟处在不同群落的情况。
1.2.2 数据收集 样地设置为20 m×20 m,并采用相邻格子法,测定、记录所有乔木的胸径、树高等,同时在样方中设4个5 m×5 m的小样方,在其中设置草本1 m×1 m样方,调查灌木(包括直径 < 2 cm的乔木)、草本(类植物)和层间植物的种类、株数(或多度)及生长状况,样地基本情况如表1。
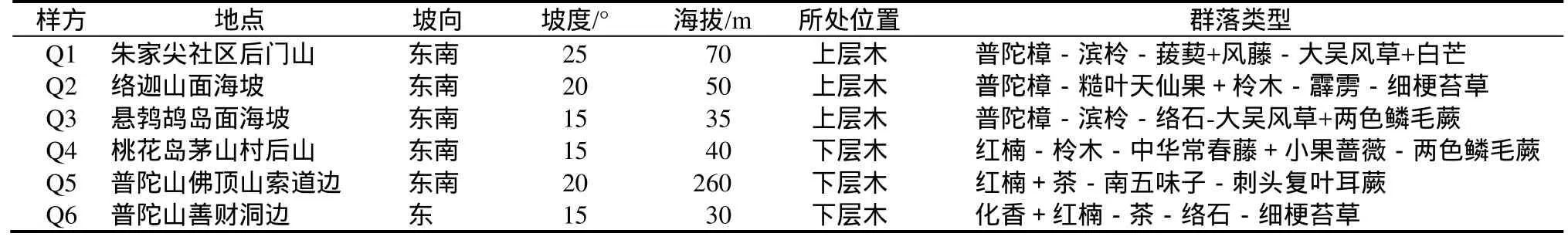
表1 调查群落生境与植被类型Table 1 The habitats and vegetation types of investigated communities
1.2.3 数据统计 重要值计算公式:
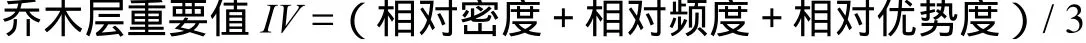
群落物种多样性的测定以重要值作为多样性指数的测度[3~4]依据,选用以下几种常用测度方法:
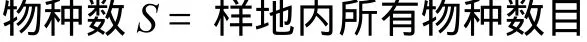
Shannon Wiener指数[5]:
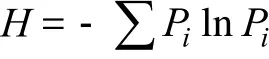
Pielou均匀度指数[3]:

Simpson 指数[6]:
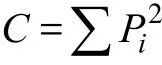
式中,Pi为第i种的个体数ni占总个体数N的比例,即Pi=ni/N。
2 结果与分析
2.1 普陀樟天然群落组成
群落中植物种类组成是形成一个群落的基础,它是群落的基本特征[5]。组成群落的植物种类多少是由群落间物种关系和环境条件共同决定的,在大区域内反映出热量和水分条件的优劣,在群落内反映了物种竞争关系和植物对所处生境的适应性,可以定量地表现为物种多样性指数的大小。
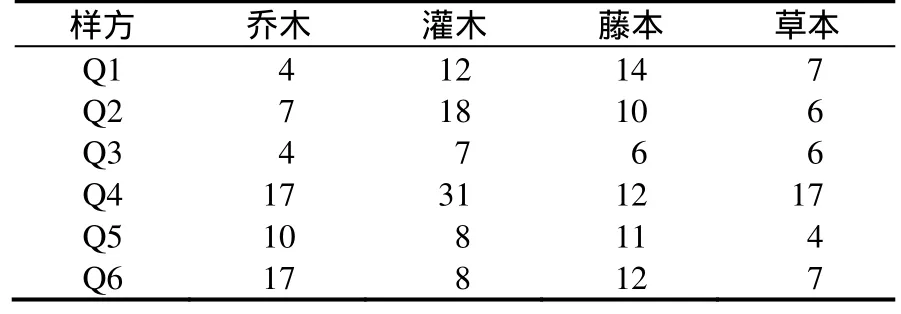
表2 调查群落物种组成Table 2 Species of investigated communities
调查表明,被调查的6个群落中与普陀樟混生的植物种类有46科72属91种。其中,乔木共有28种,灌木和藤本共有45种,草本共有18种。其中Q4群落中物种最为丰富,基本包含了所有的灌木、藤本和草本种类。普陀樟天然群落中樟科有5种:普陀樟(Cinnamonum japonicumvar.chenii)、香樟(C. camphora)、舟山新木姜子(Neolitsea sericea)、豹皮樟(Litsea coreanavar.sinensis)、红楠(Machilus thunbergii);豆科(Leguminosae):合欢(Albizia julibrissin)、山合欢(A. kalkora)、黄檀(Dalbergia hupeana);冬青科(Aquifoliaceae):铁冬青(I. rotunda)、冬青(I. purpurea)、全缘冬青(I. integra);榆科(Ulmaceae):榔榆(Ulmus parvifolia)、沙朴(Celtis tetrandra);桑科(Moraceae):构棘(Cudrania cochinchinensis)、桑(Morus alba);壳斗科(Fagaceae):青冈(Cyclobalanopsis glauca)、赤皮青冈(C. gilva);棕榈科(Palmae):棕榈(Trachycarpus fortunei);芸香科(Rutaceae):椿叶花椒(Zanthoxylum ailanthoides);卫矛科(Celastraceae):白杜(Euonymus maackii),松科(Pinaceae):黑松(Pinus thunbergii);山矾科(Symplocaceae):四川山矾(Symplocos setchuensis);山茶科(Theaceae):山茶(Camellia japonica);漆树科(Anacardiaceae):黄连木(Pistacia chinensis);金缕梅科(Hamamelidaceae):檵木(Loropetalum chinense);胡桃科(Juglandaceae):化香(Platycarya strobilacea);大戟科(Euphorbiaceae):日本野桐(Mallotus japonicus);大风子科(Flacourtiaceae):柞木(Xylosma japonica)。从伴生乔木树种出现概率上来看,以红楠最高,几乎每个样地中都有分布,沙朴、椿叶花椒、化香则出现50%以上的样地中,而其他都是偶尔出现。灌木共有33种,主要有柘(Cudrania tricuspidata)、柞木(Xylosma japonica)、糙叶天仙果(Ficus erectavar.beecheyana)、毛瑞香(Daphne odoravar.atrocaulis)、厚叶石斑木(Raphiolepis umbellata)、胡颓子(Elaeagnus pungens)、日本野桐、算盘子(Glochidion puberum)、海桐(Pittosporum tobira)、滨柃(Eurya emarginata)、糙叶天仙果、茶(Camellia sinensis)、山茶(Camellia japonica)、柃木(Eurya japonica)、朱砂根(Ardisia crenata)、檵木、映山红(Rhododendron simsii)、栀子(Gardenia jasminoides)、紫金牛(Ardisia japonica)。藤本共有14种,有南五味子(Kadsura longipedunculata)、络石(Trachelospermum jasminoides)、中华常春藤(Hedera nepalensis)、薜荔(Ficus pumila)、菝葜(Smilax china)、小果蔷薇(Rosa cymosa)、清风藤(Sabia japonica)、千金藤(Stephania japonica)、鸡屎藤(Paederia scanden)、海刀豆(Canavalia lineata)、木防己(Cocculus orbiculatus)、硕苞蔷薇(Rosa bracteata)、海金沙(Spora Lygodii)、风藤(Piper kadsura)。草本共有18种,主要有麦冬(Ophitopogin japonicum)、山麦冬(Liriope spicata)、单头紫菀(Aster turbinatus)、异盖鳞毛蕨(Dryopteris decipiens)、假红盖鳞鳞毛蕨(Dryopteris paraerythrosora)、两色鳞毛蕨(Dryopteris bissetiana)、三叶青(Tetrastigma hemsleyanum)、江南星蕨(Microsorium henryi)、刺头复叶耳蕨(Arachniodes exilis)、荩草(Arthraxon hispidus)、大吴风草(Farfugium japonicum)、求米草(Oplismenus undulatifolius)、紫花堇菜(Viola grypoceras)、疏花野青茅(Deyeuxia arundinaceaevar.laxiflora)、细梗苔草(Carex teinogyn)。
从树种组成来看,乔木中常绿树种和落叶树种各占一半,其中热带分布具有泛热带、热带亚洲分布类型较多,温带分布以北温带、东亚与北美间断分布及东亚(中国—日本)分布类型居多,其中有许多为珍贵用材树种,如红楠、黄檀、铁冬青、香樟、青冈等。就科的数目和科内所含种的数目而言,超过5种的科是,蔷薇科7种,山茶科6种,含2 ~ 5种植物的科18个,所占比例为39.13%,根据属级统计,群落中植物共计72属,没有超过5种的属,含2 ~ 5种的属共13属,占到了总属数的18.05%,仅有1种植物的属最多,达59属,总体来看,普陀樟天然群落内的优科不明显,种类主要集中在含1种的科内,在各属中的分布也是相对分散的,这种分散分布从侧面反映了科和属的组成的多样性,集中反映了群落的多样性、丰富性和复杂性。
2.2 普陀樟天然群落成层性
普陀樟在两类群落中,在不同类型的群落组成各自不同,结构也各自不同。在Q1、Q2、Q3天然群落中,普陀樟做为上层木出现,其生长茂盛,林相整齐,林冠开展,郁闭度达到99%以上。其伴生树种仅有合欢、椿叶花椒、沙朴、红楠、豹皮樟,主要为常绿阔叶林,灌木层和藤本层受上层高大阳性树木的影响,较为稀疏。主要灌木种类有柘、糙叶天仙果、毛瑞香、厚叶石斑木、胡颓子、日本野桐、算盘子、海桐、槟柃等,还有合欢、椿叶花椒、红楠、豹皮樟等树种的幼树、幼苗,尤其是Q3群落中,豹皮樟、普陀樟幼树、幼苗数量较多。藤本主要有千金藤、鸡矢藤、海刀豆、中华常春藤、木防己、硕苞蔷薇、海金沙、菝葜、风藤、络石等,与另外一组3个群落种类类似,数量较少。草本层受上层乔木层和灌木层影响,十分稀疏,草本主要有白茅、山麦冬、细梗苔草、白英等,还有两色鳞毛蕨等蕨类。
在Q4、Q5、Q6天然群落中,普陀樟做为下层木出现,属常绿、落叶阔叶混交林 ,其中Q4天然群落中普陀樟数量较少。该组3个群落内部结构分化也明显,可分为乔木层、灌木层、藤本层、草本层4个层次。乔木层以阳性树种和中性树种为主,有部分分层现象,阳性树种较为高大,中性树种有时在阳性乔木树种下方,树冠通常不大。此类群落中皮樟、普陀樟幼树、幼苗数量较多,证明天然更新能力较强,数量上比红楠为多。灌木和藤本相对较多,主要灌木种类有糙叶天仙果、茶、柃木、蚊母树、胡颓子、日本野桐、海桐、朱砂根、茶、檵木、山茶、映山红、胡颓子、毛瑞香、栀子、紫金牛等。主要藤本有南五味子、络石、中华常春藤、络石、霹雳、菝葜、小果蔷薇、菝葜、风藤等。草本层通常较为稀疏,主要草本有细梗苔草山麦冬、三叶青、疏花野青茅、大吴风草、山麦冬等,还有刺头复叶耳蕨、江南星蕨、两色鳞毛蕨等蕨类。
2.3 普陀樟天然群落乔木层主要数量
相对密度、相对优势度、相对频度和重要值等都是能够反映群落物种结构的数量指标,其中重要值表示一个种的优势程度,反映了该种群在群落中相对重要性的一个综合指标和对所处群落的适应程度。6个不同群落乔木层主要树种种类、相对密度、相对优势度、相对频度和重要值等指标见表3。
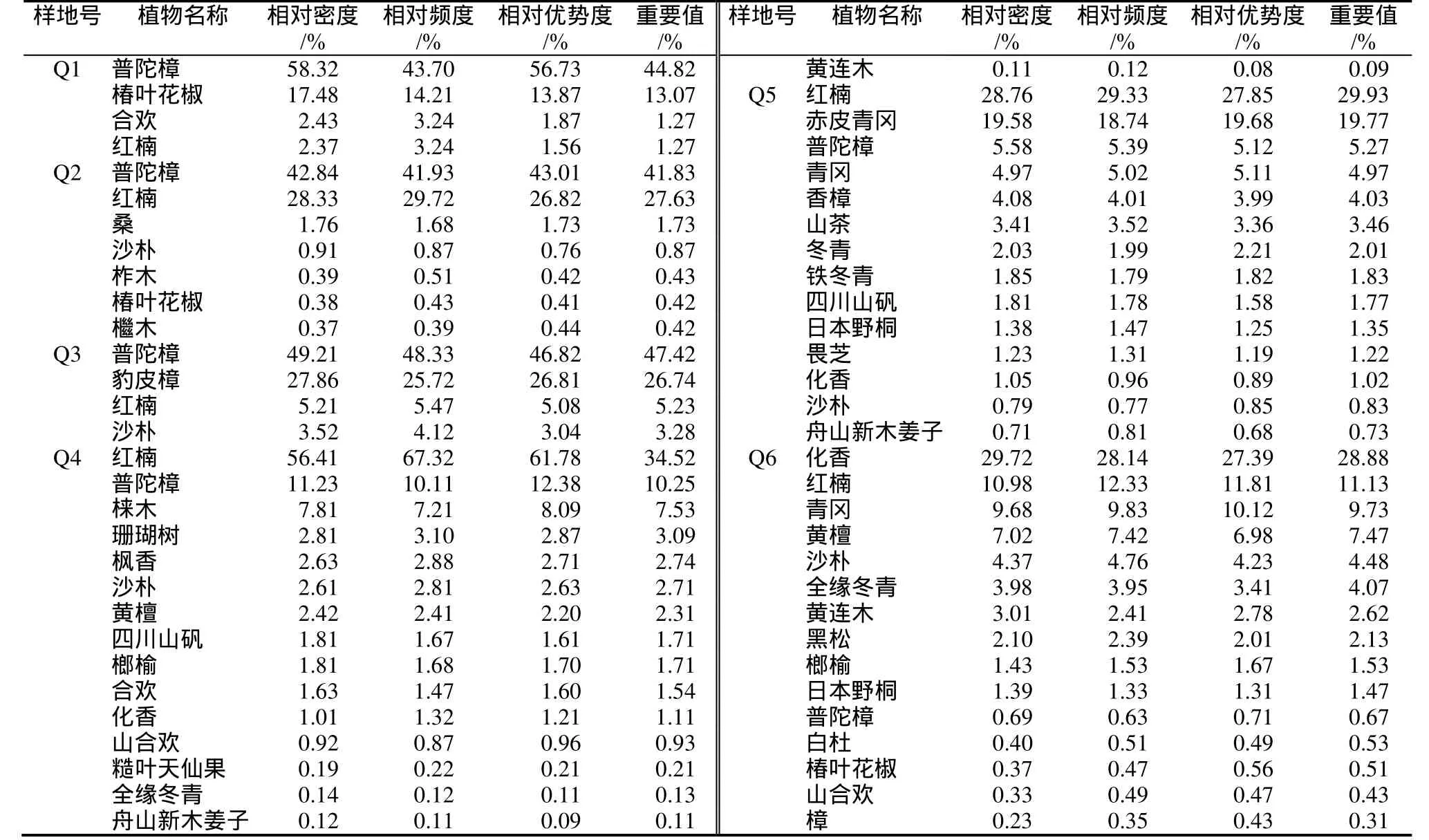
表3 普陀樟天然群落乔木层树种数量特征Table 3 The quantitative characteristics of arbor layer species in six C. japonicum var. chenii natural forest
对比表1可发现,普陀樟对环境条件要求较高,生境一旦出现人为干扰,产生一定的破坏,致其在群落中的重要值也随之下降。Q4群落所在地人为干扰最大,故此其普陀樟重要值也下降为最小。
2.4 普陀樟天然林群落乔木层物种多样性指数分析
物种多样性反映了生物群落在组成、结构、功能和动态等方面的异质性[6],体现了群落结构类型、组织水平、发展阶段、稳定程度和生境差异[7~8]。从普陀樟各天然林群落的乔木层多样性比较中可以看出(表4),6个普陀樟天然林群落乔木层中,Q4群落Simpson多样性指数、Shannon指数最高,Q5均匀度指数最高,更新能力较强,Q5群落Simpson多样性指数、Shannon指数最底,Q4群落均匀度指数最低。
一般认为当同时满足Simpson指数、Shannon指数、均匀度指数均较高时,此时物种对生态的适应性最好,而Simpson指数和Shannon指数较高、均匀度指数一般时,即使物种在群落中占优势,但对生态的适应性并不是最好。Q1、Q2、Q3群落中普陀樟虽然占优势,但并不适应该群落所在生态生境,且Q2、Q5群落处于悬水小岛,人为干扰极少,并不充分体现普陀樟的生态适应性。作为下层木出现时,群落的Simpson指数、Shannon指数、均匀度指数比作为上层木出现的组普遍较高,说明普陀樟对多样性高、立地条件较好的群落的生态适应性反而不好,调查时发现作为下层木出现的组更新数量较多的现象,也证明了这一判断。
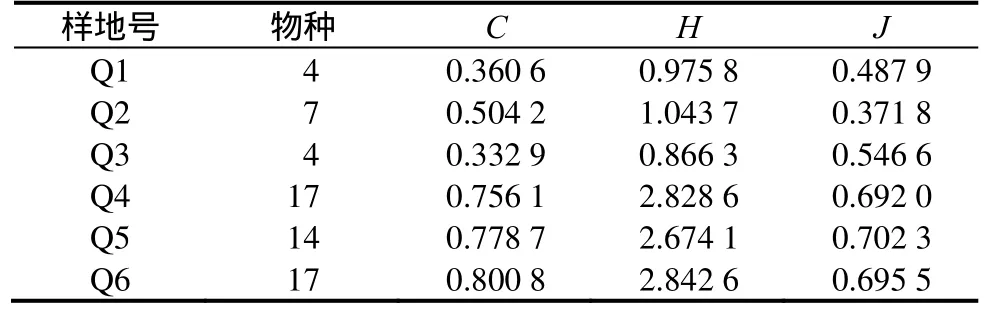
表4 普陀樟天然群落乔木层物种的多样性指数Table 4 The diversity index of arbor layer in six C. japonicum var. chenii natural forests
2.4 普陀樟天然林群落乔木层物种群种间联结性
通过对普陀樟天然林群落乔木层物种群种间联结性研究发现,普陀樟除红楠外,与乔木层其他植物的种间联结性不显著,说明普陀樟与乔本层主要树种之间不存在关联性,说明普陀樟仅和红楠对环境条件具有相同或相似的需求与适应,但是从天然分布的对比来看,红楠对环境条件适应要远高于普陀樟。普陀樟对环境条件要求较高,生境一旦出现人为干扰,产生一定的破坏,致其在群落中的重要值也随之下降。
3 结论与讨论
3.1 普陀樟分布和群落组成
普陀樟为上层木的群落的组属于常绿阔叶林,为下层木的群落的组则多是落叶树种占优势的常绿落叶阔叶混交林。在对其全面踏查时发现,普陀樟为下层木多数在常绿落叶阔叶混交林中零星分布,大乔木较少。同时从树种组成上来看,其中热带分布和温带分布多有偏亚热带地带性植物的特征。就普陀樟为上层木的群落组科属级统计看出,群落中植物单科单属种植物占主体,但数量不多。就总体的科属级统计看出,群落中植物单科单属种植物较多,反映了所在不同类型群落的多样性和复杂性。
3.2 普陀樟天然林群落乔木层重要值分析
普陀樟作为下层木在天然群落出现时,重要值普遍不高。普陀樟是中性树种,生长缓慢,多难以在正常的群落竞争中处于优势,个别如风大(Q1)、悬水小岛(Q2、Q3)等困难环境中,由于其他乔木难以生存,反而能够占有一定的优势或是成为上层木,而其一旦成为上层木,就会形成其生长茂盛,林相整齐,郁闭度极高的林相,影响其他植物的更新,从而成为一种极为特殊的顶级群落模式,同时也说明了其在特殊条件下的物种竞争力。
3.2 普陀樟天然林群落乔木层物种多样性指数分析
从6组群落数据来看,普陀樟天然林群落多样性指数分组与按照普陀樟在群落中的生态位类型划分的组类似,普陀樟为上层木的群落组,这一类普陀樟虽然占优势,但由于所处环境特殊并不适应该群落所在生态生境,所在条件特殊且人为干扰不多,并不充分体现普陀樟的生态适应性。普陀樟为伴生的群落组,群落多样性指数都较高,说明普陀樟在群落生态多样性好的条件下反而难以成为优势种,也侧面反映其对此类立地条件中竞争力不强。
3.3 普陀樟天然林保护
普陀樟天然更新能力较强,甚至在同一群落中超过广泛分布的红楠。从整个调查情况可以看出,虽然普陀樟天然分布范围不大,但林下天然更新较好,未来可以采取适宜的措施,加强对现有天然林资源的保护和利用。在遗传资源收集、保存和培育的基础上,针对当前低质天然林现状,可通过补植造林等手段,改善林相,同时加强培育普陀樟苗木,促进普陀樟资源开发。
[1]俞慈英,陈叶平,袁燕飞,等. 舟山海岛普陀樟等3种特有树种种质资源清查[J]. 浙江林学院学报,2007,24(4):413-417.
[2]陈叶平,缪玲霞,袁燕飞,等. 普陀樟天然林的调查研究[J]. 现代农业科技,2009(9):32-33
[3]金则新. 浙江天台山甜槠群落物种多样性研究[J]. 云南植物研究,1999,21(3):296-302.
[4]贺金生,陈伟烈,李凌浩. 中国亚热带东部常绿阔叶林主要类型的群落多样性特征[J]. 植物生态学报,1998,22(4):303-311.
[5]王春玲,郭泉水,谭德远,等. 准噶尔盆地东南缘不同生境条件下梭梭群落结构特征研究[J]. 应用生态学报,2005,16(7):1224-1229.
[6]茹文明,张金屯,毕润成,等. 山西霍山森林群落林下物种多样性研究[J]. 生态学杂志,2005,24(10):1139-1142.
[7]中国森林编委会. 中国森林[M]. 北京:中国林业出版社,1978
[8]郭其强,张文辉,何景峰,等. 黄龙山不同白桦林群落结构特征研究[J]. 西北植物学报,2007,27(1):132-138.
猜你喜欢
建筑与预算(2022年6期)2022-07-08 13:59:18
少先队活动(2021年9期)2021-11-05 07:31:14
山西林业(2021年2期)2021-07-21 07:29:28
农村百事通(2020年21期)2020-11-23 02:04:24
小学生作文(低年级适用)(2019年11期)2019-11-29 02:00:56
绿色中国(2019年19期)2019-11-26 07:13:16
绿色中国(2019年19期)2019-11-26 07:13:16
科技创新与品牌(2015年2期)2015-03-18 17:47:15
建筑与预算(2014年5期)2014-04-11 07:29:28
河南科技(2014年16期)2014-02-27 14:13:33
