
BrdU免疫组织化学染色方法的改进(漂片法)
2012-07-28 10:16:42郭晶晶单立冬
中国医药导报 2012年5期
郭晶晶 单立冬 吴 钢
1.上海复旦大学附属华山医院宝山分院普外科,上海 200431;2.苏州大学医学部基础医学与生物科学学院神经生物学与医学心理学系,江苏 苏州 215123;3.上海复旦大学附属华山医院普外科,上海 200040
5-溴-2-脱氧尿嘧啶核苷(5-bromo-2-deoxyuridine,BrdU)是胸腺嘧啶核苷的类似物,在细胞周期的S期掺入到增殖或分裂细胞的核内,只要细胞不消亡,BrdU将永久地存留在胞核DNA中,且在体或离体都能通过免疫组织化学的方法检测到,可用于标记BrdU暴露期间进行DNA合成的细胞[1]。故BrdU阳性细胞可被看作是具有增殖活性的细胞。
1 材料与方法
1.1 材料
动物:SD大鼠,雌雄不拘,体重220~230 g,苏州大学实验动物中心提供。试剂:BrdU(Roche Diagnostic Co.)。
1.2 分组
踏转轮运动对成年大鼠海马齿状回细胞增殖的影响。实验分两组,①踏转轮运动组:转轮速度为10 m/min,每天训练2 次(09:00,15:00),每次训练 30 min,连续 6 d;②对照组:动物置于转轮中,速度为0。放置次数与时间同运动组。在染色过程中每一组又分为三个亚组,分别为加入羊血清组、胰酶抑制剂组和牛血清白蛋白组。
1.3 BrdU标记
踏转轮运动的第4天开始给药,于每次运动前1 h腹腔注射 BrdU(50 mg/kg,美国 ROCHE 公司),2 次/d,连续 3 d。 末次给药后24 h处死动物,从前囟后3.3~5.1 mm作连续冰冻切片,进行BrdU染色。
1.4 BrdU免疫组织化学
切片经2 mol/L HCl(室温30 min)变性,用0.01 mol/L PBS充分漂洗后转入0.1%胰酶中(37℃15 min)消化后,0.01 mol/L PBS漂洗(5 min×3次),分别加入羊血清组、胰酶抑制剂组和牛血清白蛋白组,室温下30 min,然后加入小鼠抗BrdU单克隆抗体(1∶200,Biosource)室温孵育 16h,0.01mol/L PBS 漂洗(5min×3次),转入羊抗小鼠二抗(1∶200,Vector)室温孵育 2 h,再用 0.01 mol/L PBS漂洗(5 min×3次),移入亲和素生物素试剂(avidin-biotin complex,ABC,1∶200)室温孵育 2 h,最后用含0.03%H2O2的0.05%DAB 显色 3~5 min,0.01 mol/L PBS终止显色。常规脱水、透明、封片。镜检BrdU阳性细胞计数。
1.5 统计学方法
随机选取每只大鼠位置相同的5张不连续切片,光学显微镜下 (×200)计数每张切片双侧海马齿状回颗粒细胞层(granular cell layer,GCL)免疫反应阳性细胞数。阳性细胞的均数作为统计数据,以均数±标准差()表示。用SPSS 10.0软件作方差分析(one-way ANOVA)检测组间差异性。
2 结果
2.1 踏转轮运动对成年大鼠齿状回细胞增殖的作用
经过6 d的踏转轮运动训练后,各组BrdU阳性细胞见图1。免疫组织化学结果显示,海马齿状回BrdU阳性细胞数,对照组为(22.55±1.91)个/切片,踏转轮组为(57.89±1.90)个/切片,两组相比差异有高度统计学意义(P<0.001)。由此可见,踏转轮运动可增加BrdU阳性细胞,促进齿状回细胞增殖。
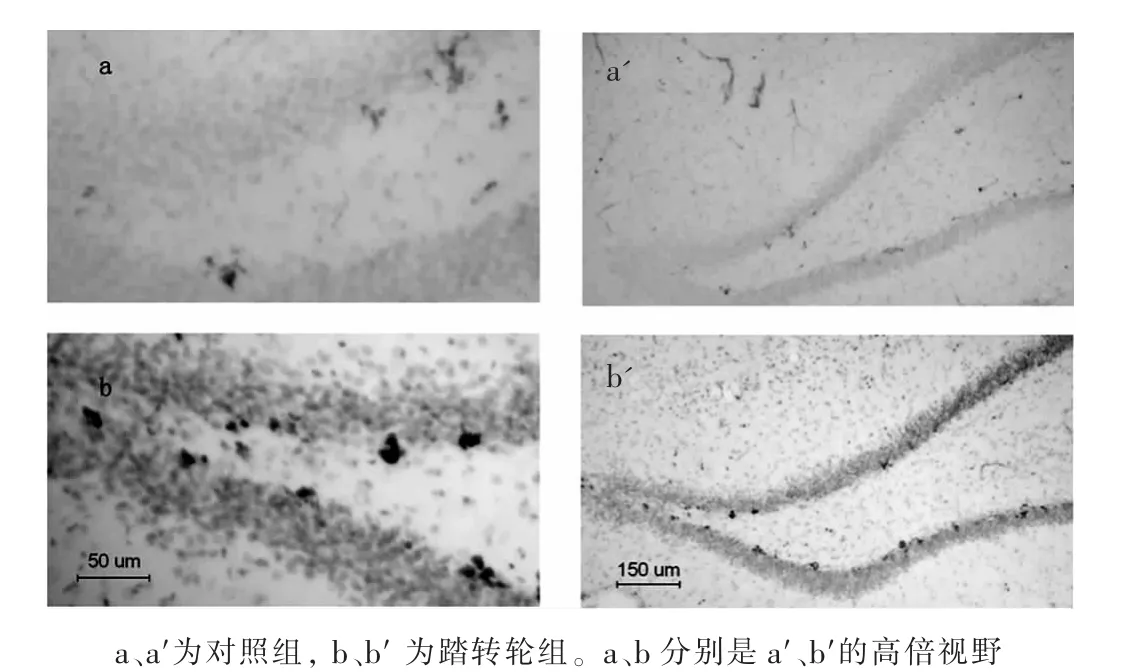
图1 踏转轮运动6 d后海马齿状回BrdU阳性细胞数
2.2 不同的封闭液对组织片染色效果的影响
末次给药后24 h处死动物,从前囟后3.3~5.1 mm作连续冰冻切片,将切片随机分成三组。在染色过程中,这三组分别加入羊血清、胰酶抑制剂和牛血清白蛋白作为封闭液和终止胰酶作用的抑制剂。各组BrdU阳性细胞如图2所示。免疫组织化学结果显示,三种方法均能使BrdU着色,但各组的本底颜色和非特异性着色程度不同:羊血清组中虽然阳性细胞与非特异性着色细胞有明显区别,但非特异性着色细胞显色度较深,且遍布整个视野。胰酶抑制剂组阳性细胞与非特异性着色细胞也有明显差异,而且非特异性着色细胞显色度较浅,整个本底相对较为干净、清爽。牛血清白蛋白组几乎见不到非特异性着色细胞,本底颜色非常淡。
除了本底颜色和非特异着色的观察外,笔者还重点观察了组织切片的破损情况。实验结果显示:牛血清白蛋白和胰酶抑制剂组无切片破损,而羊血清组有少部分脑片破损。细胞计数显示:羊血清组为(23.22±2.38)个/切片,胰酶抑制剂组为(22.54±2.12)个/切片,牛血清白蛋白组为(23.14±3.12)个/切片,各组间差异无统计学意义(P>0.05)。
3 讨论

图2 不同封闭液对BrdU免疫组织化学染色效果的影响
自从20世纪末神经干细胞发现以来,其目前已经成为神经科学研究的热点之一。而研究在体或移植后的神经干细胞的增殖、迁移、分化中常用的标记新生细胞的方法有BrdU、3H、Ki-67、 PCNA(proliferating cell nuclear antige)[2-3],前两者在细胞增殖时整合入细胞以后,即使在细胞静止期仍能表达;后两者是细胞增殖过程中表达的内源性蛋白,当细胞处于静止期时,它们并不表达。因此,BrdU和3H是常用的追踪细胞的标记物,而BrdU的检测又较3H更为方便、简单,所以BrdU是最为常用的标记细胞增殖的标记物。
丰富环境对神经发生有影响,而在各种环境因素中,运动对其影响较为显著[4-5]。本课题的前期研究建立了踏转轮运动的模型[6],并发现运动对神经发生的作用具有强度依赖性,这早于国内外的相关报道[7-8]。故本课题选用运动作为促进细胞增殖的方法,观察了不同的染色方法对切片及结果的影响。
组织切片BrdU染色时,常用的方法有贴片法和漂片法。漂片法与贴片法相比有一定的优点:前者能使抗体从两侧渗透,更有利于抗体与抗原充分反应,相应的在洗片时,也能使未结合的抗体充分漂洗干净;漂片法比贴片法所使用的抗体数量少,而且必要时抗体还可以回收,重复利用。标本量大时,漂片法比贴片法更省时、省力。但是,用漂片法进行BrdU染色时,由于在加入抗体前要用盐酸和胰酶对脑片进行预处理,经过处理(尤其是胰酶处理)后,进行染色的脑片极易破损,从而导致整个染色过程无法正常完成,或最终所得到的脑片极少。因此,在本实验中针对胰酶的消化作用,笔者分别选用了与二抗同源的羊血清、牛血清的主要成分白蛋白(牛血清白蛋白)及胰酶抑制剂来抑制(或终止)胰酶的消化作用。
羊血清是免疫组织化学中常用的封闭液,但是,在本实验中并未取得预期的成果,虽然羊血清能终止胰酶的消化作用,但并未能有效地封闭非特异性染色。其原因目前仍不清楚,有待进一步研究。胰酶抑制剂(取自火鸡蛋清)组和牛血清白蛋白组非特异性染色较少,尤其是牛血清白蛋白组。其原因可能是二者的成分单一,纯度较高。
[1]Brent AR,Weiss S.Generation of neurons and astrocytes from isolated cells of the adult mammalian central nervous system [J].Science,1992,255(5052):1707-1710.
[2]Cameron HA,Mckay RD.Adult neurogenesis produces a large pool of new granule cells in the dentate gyrus[J].J Comp Neurol,2001,435(4):406-417.
[3]Kee N,Sivalingam S,Boonstra R,et al.The utility of Ki-67 and BrdU as proliferative markers of adult neurogenesis[J].J Neurosci Methods,2002,115:97-105.
[4]Praag van H,Kempermann G,Gage FH.Running increases cell proliferation and neurogenesis in the adult mouse dentate gyrus[J].Nat Neurosci,1999,2(3):266-270.
[5]Zhao C,Deng W,Gage FH.Mechanisms and functional implications of adult neurogenesis[J].Cell,2008,132(4):645-660.
[6]Xu WP,Shan LD,Gong S,et al.Forced running enhances neurogenesis in the hippocampal dentate gyrus of adult rats and improves learning ability[J].Acta Physiologica,2006,58(5):415-420.
[7]Kronenberg G,Bick-Sander A,Bunk E,et al.Physical exercise prevents age-related decline in precursor cell activity in the mouse dentate gyrus[J].Neurobiol Aging,2006,27(10):1505-1513.
[8]娄淑杰,刁玮,陈佩杰.运动频率和递增强度运动对大鼠海马神经发生的影响[J].体育科学,2010,30(1):66-69.
猜你喜欢
北方药学(2022年3期)2022-09-06 00:22:06
——“AABC”和“无X无X”式词语
小学生学习指导(中年级)(2021年3期)2021-04-06 09:12:02
少儿美术(快乐历史地理)(2020年8期)2021-01-04 01:22:44
福建畜牧兽医(2018年1期)2018-03-01 08:28:44
中西医结合心血管病电子杂志(2016年36期)2017-07-05 14:30:28
兰台世界(2017年12期)2017-06-22 14:17:25
中国组织化学与细胞化学杂志(2017年1期)2017-06-15 20:27:43
中国组织化学与细胞化学杂志(2016年3期)2016-02-27 11:15:28
中国医学科学院学报(2015年5期)2015-03-01 04:03:42
中国兽药杂志(2015年12期)2015-01-25 10:06:12
