
产异淀粉酶嗜热菌选育及产酶条件的研究
2012-07-10 08:48:40胡先望严晓娟
湖南农业科学 2012年1期
胡先望 ,陈 朋 ,梁 宁 ,王 雄 ,严晓娟
(1.甘肃省商业科技研究所,甘肃 兰州 730010;2.甘肃农业大学动物医学院,甘肃 兰州 730070)
异淀粉酶在生化分类上属于淀粉水解酶类(EC3.2.1.68)。它可以从寡聚糖的非还原末端水解切开α-1,6糖苷键,释放出葡萄糖,可广泛应用于淀粉糖浆、啤酒和酒精等的生产。近年来,以淀粉为原料利用酶技术开发出的低聚异麦芽糖因其安全无毒、理化性质独特等优良性质而备受关注[1-2]。异淀粉酶在高浓度的葡萄糖环境中也能将游离葡萄糖残基通过酶催化逆反应将葡萄糖苷转移到另一底物形成α-1,6糖苷键,从而得到非发酵性的低聚糖[3]。以淀粉为原料,经过α-淀粉酶、β-淀粉酶和异淀粉酶(或α-葡萄糖苷酶)的协同作用,可生产低聚异麦芽糖或糖醇,其转化率一般为55%~60%[4]。我国工业用异淀粉酶主要来自进口,近年来国内朱学军[5]、王弋博[6]、夏静[7]、郭宏文等[8-9]分别进行了产异淀粉酶菌种的选育工作,其他关于嗜热异淀粉酶的研究报道则较少。笔者以嗜热脂肪芽孢杆菌为原始菌种,进行诱变及优势菌株选育,并对筛选出的突变菌株的产酶性质做了初步探讨,旨在获得耐高温、活性较高、稳定性较好的产酶菌株。
1 材料与方法
1.1 材料
1.1.1 菌 种 嗜热脂肪芽孢杆菌(Bacillus stearothermophilus,菌种编号 1.1865),购于中国科学院微生物研究所,经诱变处理得到高产异淀粉酶的突变菌株UM761。
1.1.2 仪器与试剂 仪器:TU-1800PC紫外可见分光光度计(北京普析通用仪器有限责任公司)、GL-21M离心机(湖南湘仪离心机仪器有限公司)、MF99-3自动核酸蛋白分离层析检测仪(上海沪西分析仪器厂)、LDZM-75型立式智能压力蒸汽灭菌器(上海申安医疗器械厂)、5 L全自动发酵罐(日本B.E.Marubsishi)等。试剂:琼脂、蛋白胨、牛肉浸膏(北京双旋微生物培养基厂)、氯化钠、磷酸二氢钠、醋酸钠、冰乙酸、盐酸、氢氧化钠(上海试剂三厂)、可溶性淀粉(北京化工试剂厂)、硫酸铵(西安试剂厂)、3,5—二硝基水杨酸(DNS)、硝基胍、牛血清白蛋白(中科院上海生化研究所)、酵母粉(兰州金宝瑞胶囊有限责任公司)。
1.2 方法
1.2.1 菌种培养 将嗜热脂肪芽孢杆初始菌接种于LB营养肉汁培养基中,45℃培养24 h,取菌液1 mL测定菌密度A600。当A600≥2.0后,培养液8 000 r/min离心5 min。收集上清液,测定异淀粉酶活力。
1.2.2 菌种诱变 取培养至对数期菌液10 mL(OD600=0.569),3 000 r/min离心 15 min,弃上清,用无菌生理盐水制成菌悬液,用血球计数板在显微镜下计数(108个/mL)。取5 mL加入到含有0.5 mg/mL的亚硝基胍溶液的培养皿中,并将处理皿放在无菌磁力搅拌器,15 W、254 nm紫外灯30 cm处照射。用紫外灯照射复合处理,分别于30、40、50 min取出0.5 mL的培养液,加入0.5 mL 25%的硫代硫酸钠溶液终止反应,然后稀释涂平板,取未经NTG处理的菌液稀释涂平板做对照。将上述平板于45℃培养20 h后计数,并计算致死率。培养20 h后,挑出大、中、小菌落48个,分别培养后并测定释放酶活,从中选出酶活最高的菌株。
1.2.3 酶液体深层发酵培养及分离 将液体试管菌种接入三角瓶液体培养基中,45℃摇瓶培养24~96 h,随时观察菌种生长情况,检测菌密度和异淀粉酶活力。培养结束后,经12 000 r/min离心过滤,上清即为粗酶液。酶液用硫酸铵沉淀,收集饱和度30%~80%的沉淀,经0.1 M pH值6.2的醋酸盐缓冲液透析后,得到初步纯化的酶产品。
1.2.4 酶活力测定 α-淀粉酶活力测定[10-12]:取一定量的酶液稀释至合适的浓度,与1%的可溶性淀粉(溶于0.1 M pH值6.0的醋酸盐缓冲液中)50℃反应30 min,然后添加3,5—二硝基水杨酸显色试剂(DNS),混匀后沸水浴5 min,立即冷却。用0.1 M pH值6.0的醋酸盐缓冲液稀释至合适的体积后,测540 nm处的OD值,根据标准曲线可求得水解产生的还原糖的量。酶活力单位定义为在上述条件下每分钟水解淀粉生成1 μmol葡萄糖所需要的酶量(mL)为1 U。
异淀粉酶活力测定:取一定量的酶液稀释至合适的浓度,与1%的普鲁兰(溶于0.1 M pH值6.0的醋酸盐缓冲液中)50℃反应30 min,然后添加3,5—二硝基水杨酸显色试剂(DNS),混匀后沸水浴5 min,立即冷却。用0.1 M pH值6.0的醋酸盐缓冲液稀释至合适的体积后,测540 nm处的OD值,根据标准曲线可求得水解产生的还原糖的量。酶活力单位定义为在上述条件下每分钟水解淀粉生成1 μmol葡萄糖所需要的酶量(mL)为1 U。
2 结果与分析
2.1 菌种诱变酶活测定
初始菌株嗜热脂肪芽孢杆菌培养60~72 h,α-淀粉酶活力为1.0~1.5 U/mL,异淀粉酶活力为2.5~3.2 U/mL;经诱变筛选获得的变异菌株UM761培养 48~60 h,α-淀粉酶活力为 0.9~1.5 U/mL,异淀粉酶活力为5.5~9.6 U/mL。与初始菌株产酶活性比较,UM761酶活比初始菌株提高了2~3倍,培养时间缩短1/4,α-淀粉酶活力基本没变。
2.2 UM761菌种产酶条件初步优化
2.2.1 培养基不同碳源对产酶的影响 选择葡萄糖、麦芽糖和可溶性淀粉作为菌种培养碳源,其他培养基成份和培养条件都相同,根据培养时间测定培养液的菌密度和培养液中的酶活并与碳源进行比较,结果如图1、图2所示,葡萄糖和可溶性淀粉是较好的碳源。选择培养基葡萄糖的浓度分别为0.05%、0.1%、0.2%、0.3%、0.4%进行摇瓶培养 72 h,并测定培养液中酶活力,结果见表1,培养基中葡萄糖浓度达到0.1%时,菌种生长及培养液中异淀粉酶生成可以达到最佳状态。
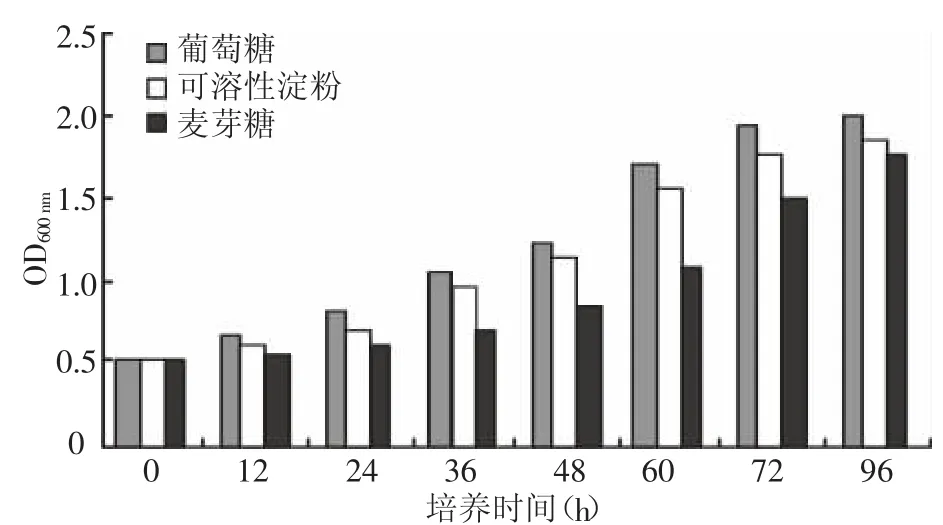
图1 不同碳源培养时间与细菌密度的关系
2.2.2 培养基不同氮源对产酶的影响 选择牛肉汁、牛肉浸膏、蛋白胨、酵母粉作为菌种培养氮源,其他基成份和培养条件都相同,根据培养时间测定培养液的菌密度和培养液中的酶活并与氮源进行比较,结果如图3、图4所示。
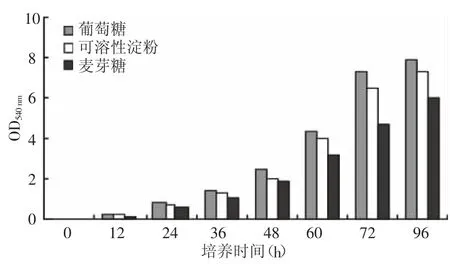
图2 不同碳源培养时间与酶活力的关系

表1 培养基中葡萄糖浓度与菌种生长的关系
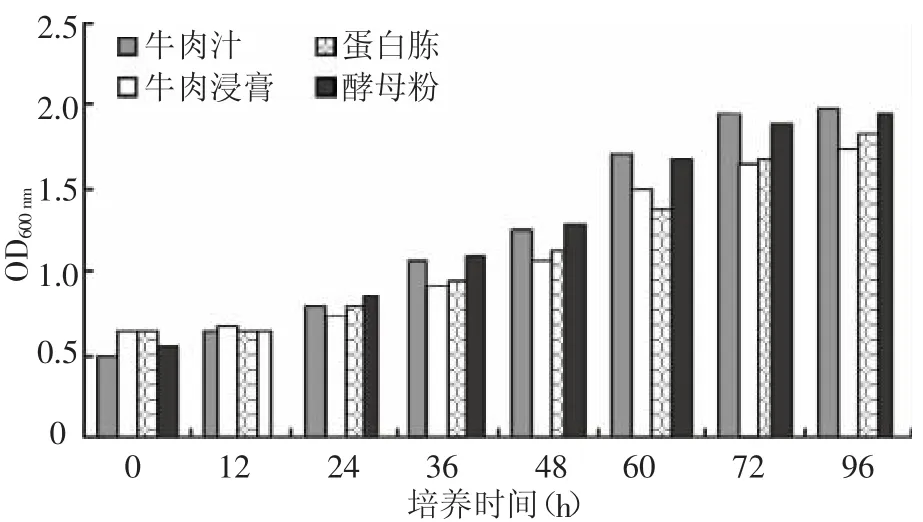
图3 不同氮源培养时间与菌密度的关系
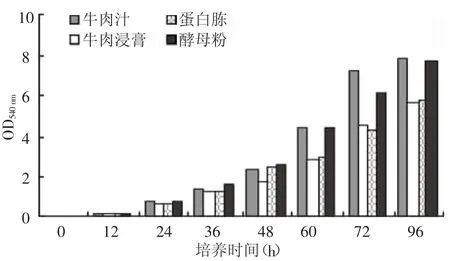
图4 不同氮源培养时间与酶活力的关系
牛肉浸膏和酵母粉都是较好的氮源,酵母粉也可以促进菌种的快速生长和繁殖,但没有促进酶分泌的作用。由于牛肉汁主要用新鲜肉制得,价格昂贵,不适宜用于实际生产,因此选用牛肉浸膏、蛋白胨和酵母粉的混合物作为菌种氮源,菌种的生产周期为72 h左右,培养液酶活力可达9.0 U单位以上。
2.2.3 培养温度对产酶活性的影响 根据以上确定的培养基,选择培养温度分别为35、40、42、44、45、48、50、55℃进行三角瓶振荡培养,其他培养条件都相同,根据培养时间测定培养液的菌密度和培养液中的酶活,结果如图5、图6所示,菌种最佳培养温度为44~46℃。

图5 不同培养温度与菌密度的关系

图6 不同培养温度与酶活力的关系
2.2.4 培养时间对产酶活性的影响 在最适培养条件下,每隔8 h测定培养液的酶活力,结果如图7所示,在培养48 h时产酶活性最高,最终确定菌种最佳培养时间为48~64 h。

图7 不同培养时间对菌株产酶活性的影响
2.2.5 培养基初始pH值对产酶活性的影响 配制不同pH值的发酵培养基,同步发酵,测定菌密度与酶活。制作培养基初始pH值与产酶的关系曲线如图8所示。由图8可知,菌株可在广泛的初始pH范围内发酵,在偏酸性近中性的环境中发酵的酶活均较高,pH值在6.2时酶活达到最高,菌种培养最佳初始pH为6.2~6.5。
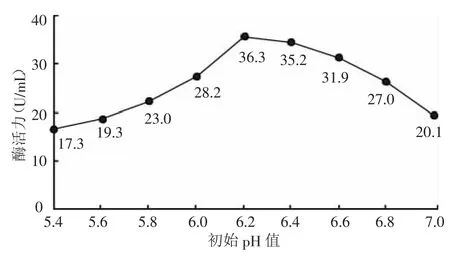
图8 不同初始pH值对产酶活性的影响
2.2.6 金属离子对产酶活性的影响 在酶反应系统中分别添加终浓度为 10 mmol的 Co2+、Mg2+、Ca2+、Mn2+、Ni2+、Fe2+、Fe3+、PO43-、HPO43-、EDTA,在最适培养条件下进行培养,以不添加金属离子酶活性为100%,计算酶的相对活性。结果如表2所示,Co2+、Mg2+、Ca2+对异淀粉酶有较强的激活作用,而Fe3+、Fe2+对酶则有较强的抑制作用,异淀粉酶在磷酸盐中很稳定。

表2 金属离子对酶活力的影响
3 讨论
微生物所产的淀粉酶在淀粉工业运用中具有举足轻重的作用,可以与α-淀粉酶、β-淀粉酶等水解酶相互协同,提高淀粉的转化率,并有效地增加出糖率。嗜热的异淀粉酶市场需求大,产品附加值高,投资回报率高,特别是嗜热异淀粉酶工程菌的开发和应用,将填补国内空白,对增加我国酶制剂行业的品种,促进国内生物酶高新技术的研发,提高淀粉的附加值,降低生产成本,增加企业的经济效益等都有重要的意义。研究选取嗜热脂肪芽孢杆菌进行产异淀粉酶菌种诱变,与原始菌株相比,诱变所获得的UM761菌株产酶效率提高了2~3倍。在此基础上,笔者对此菌株的产酶条件做了优化,最适产酶菌株培养条件为以1%葡萄糖作为碳源,牛肉膏,蛋白胨和酵母粉的混合物作为氮源,温度44~46℃,培养时间48~64 h,培养基初始pH值6.2~6.5。同时研究了金属离子对产酶活性的影响,结果表明Co2+、Mg2+、Ca2+对异淀粉酶有较强的激活作用,而Fe3+、Fe2+对异淀粉酶则有较强的抑制作用,而异淀粉酶在磷酸盐中很稳定。
[1]毕金峰,刘长江.低聚异麦芽糖的特性及其应用[J].粮油食品科技,2002,10(5):42-44.
[2]尤 新.我国低聚糖生产技术研究进展 [J].食品工业科技,2002,23(4):4-7.
[3]张应玖,朱学军,关 键,等.一种新型淀粉酶的鉴定及其产酶菌株的筛选[J].微生物学通报,2002,29(5):38-41.
[4]林亲录,符 琼,周丽君.低聚异麦芽糖制备研究进展[J].食品工业科技,2011,(2):352-354.
[5]朱学军,李吉平,孟庆繁,等.高产新型淀粉酶菌株M S511的选育及最适产酶条件 [J].吉林大学学报(理学版),2002,40(2):196-199.
[6]王弋博,李 博,李三相,等.异淀粉酶产生菌的分离纯化及生长特性[J].青海师范大学学报(自然科学版),2003,(1):82-85.
[7]夏 静,陈朝银,上官俊龙,等.一株产异淀粉酶栖热菌的生长和产酶初步研究[J].微生物学杂志,2005,25(5):107-109.
[8]郭宏文,冮 洁,邹东恢.异淀粉酶产生菌的产酶性质研究[J].食品科技,2010,35(12):17-19.
[9]郭宏文,冮 洁.异淀粉酶产生菌的筛选 [J].食品科技,2011,36(2):8-21.
[10]赵 鹏,白晓雷,韩海霞,等.赤霉素对不同温度下沙芥种子萌发特性及α-淀粉酶活性的影响 [J].华北农学报,2011,26(1):127-130.
[11]刘雅琴,乌日娜,段金华.耐酸性α-淀粉酶产生菌的发酵条件优化[J].安徽农业科学,2010,38(35):19888-19890.
[12]郭红祥,尹 钧.反义Trx s基因对小麦种子萌发中a-淀粉酶的影响[J].江西农业学报,2009,21(6):1-3.
猜你喜欢
中国粮油学报(2019年4期)2019-07-12 09:06:32
山东工业技术(2016年15期)2016-12-01 05:31:50
中国酿造(2016年12期)2016-03-01 03:08:19
中国酿造(2016年12期)2016-03-01 03:08:11
应用海洋学学报(2015年3期)2015-11-22 07:39:30
中国酿造(2014年9期)2014-03-11 20:21:03
食品工业科技(2014年15期)2014-03-11 18:17:52
食品工业科技(2014年9期)2014-03-11 18:15:55
食品工业科技(2014年9期)2014-03-11 18:15:28
食品工业科技(2014年7期)2014-03-11 18:14:49
