
鱼类对水动力环境变化的行为响应特征
2012-04-17 09:30:24严忠民
河海大学学报(自然科学版) 2012年4期
齐 亮,杨 宇,王 悦,严忠民
(1.河海大学水文水资源与水利工程科学国家重点试验室,江苏南京 210098;
2.河海大学水利水电学院,江苏南京 210098;
3.南京水利科学研究院水工水力学研究所,江苏南京 210029;
4.南京大学生命科学学院,江苏南京 210093)
目前研究鱼类在水中受到水流刺激之后的行为主要集中在2个研究领域:鱼类学领域和水力学领域。鱼类学主要有鱼类生理学和鱼类行为学2个学科。鱼类生理学是针对鱼类响应水流刺激的感受器构造及其机理进行研究,鱼类行为学主要对鱼类受到刺激后的行为进行观察,总结其行为规律。而在水力学方面,随着水利工程对鱼类种质资源影响的日益突出,对鱼类栖息地、鱼类过坝设施等相关研究也逐渐受到重视,进而作为交叉学科的生态水力学得到进一步发展,鱼类在水中的行为研究也越来越受到水力学研究者的重视。然而由于鱼类学和水力学的学科差异,以及两方面的研究者知识背景的不同存在较深的学术鸿沟。本文通过对鱼类学领域和水力学领域的鱼类行为试验的总结分析,寻找构建2个领域交叉研究桥梁的方法,为鱼类感觉阈值与水流的空间分布试验提供方法支持。
1 鱼类感觉水动力特征的生理机理
鱼类依靠侧线系统感觉栖息地的水动力特征。鱼类的侧线系统以分散在身体不同部位的神经丘为基础,在鱼类与周围水环境发生相对运动时感觉水流运动[1]。组成侧线系统的神经丘分为侧线管神经丘与体表神经丘,这2种神经丘都是由感觉细胞来传递水流产生的刺激,但由于分布位置不同以及感觉细胞数量和形态上的差异[2],导致了2种神经丘有不同的功用。
水流运动对神经丘的刺激由感觉细胞来实现。由于感觉细胞的感觉毛对位移敏感,感觉细胞相当于位移传感器,其反应的强度由静纤毛偏移的方向和幅度来确定[3]。当水流运动使感受器的感觉顶发生倾斜时,位于下部的感觉细胞就发出冲动:若静毛倒向动毛一侧,细胞产生去极化(inhibitory),神经纤维发放冲动频率增加;若动毛倒向静毛一侧,细胞产生超极化(excitatory),神经纤维发放冲动频率减少[4]。这些变化通过感觉神经纤维传递到神经中枢,于是鱼类就对水流产生了感觉。图1为感觉细胞作用示意图。另外,在单个的体表神经丘和侧线管神经丘中,相邻的2个感觉细胞其动纤毛位置是相反的,也就是说感觉顶的倾斜会导致相邻的感觉细胞产生相反的极化反应[5]。

图1 感觉细胞及其方向敏感性示意图Fig.1 Schematic diagram of a hair cell and its directional sensitivity

图2 侧线及侧线管神经丘示意图Fig.2 Schematic diagram of lateral line and canal neuromasts
侧线管神经丘位于鱼类的侧线中。侧线是鱼类和两栖类特有的感觉器官,多分布在头部以及身体的两侧。侧线管内充满黏液,侧线管神经丘浸在黏液中。鱼类发育的初期,侧线管神经丘往往分散排列,此时感觉细胞的感觉顶露于体表。随着个体的发育成长,侧线管神经丘便会逐渐沉入皮下成为短沟,然后彼此联通形成封闭的长管,而且完全与皮肤分开,仅以一个个小孔与外界相通(图2)。小孔之间压力不同会导致侧线管内液体运动,触动感觉毛,对感觉细胞产生刺激,因而侧线管神经丘能够对压力产生感觉。因为当相邻小孔之间存在流速梯度时,会产生压力差,所以侧线管神经丘可以作为流速梯度感受器。此外,侧线管神经丘对空间内的非均匀流有反应,如小范围震动产生的非均匀流和从静止鱼类身边通过的物体产生的非均匀流[6]。由神经丘组成的侧线系统能够接受不同方向的各种刺激,使得鱼类能够获取周围水环境中足够的信息,从而对水流产生感觉。
体表神经丘分散在鱼类身体表面,能够探测流经鱼类体表的水流强度和方向,以及水流中切应力区域,使鱼类表现出趋流性和其他游泳行为[7-8],这样体表神经丘就能获得空间内均匀流或非均匀流的信息。在不同水流条件下,体表神经丘和侧线管神经丘对于水力刺激的反应不同:体表神经丘在静水中对周边水体的震动较为敏感,在单向水流中对震动几乎没有反应;侧线管神经丘对震动的反应则不受水流的影响,在静水或动水中都对震动有反应。
2 鱼类对流速变化的响应
2.1 鱼类对流速感觉的试验
鱼类的神经丘功用不同,体表神经丘能够作为速度感受器,探测鱼类身体周围水流速度的强度和方向,Voigt等[7]的研究证明了这点。淡水鳗鱼喜欢在夜晚活动,在微光条件下视觉难以发挥作用,所以主要依赖侧线系统感受水流,从而调整游泳行为。因而研究者以淡水鳗鱼为目标种,首先对其实施手术,将目标种中脑和端脑之间的脑组织切除,露出侧线神经纤维的根部,再将神经纤维与充满NaCl(0.4mol/L)的微电极管相连,神经信号经过放大可在示波器上观察,神经信号的波形、振幅和峰电位间距是判断其是否产生冲动的标准。把目标种处理过后,放入水槽(100cm×25cm×10cm)中,如图3所示。水槽由带隔板的进水段(图3中1段)、有水管阵列的上下游段(图3中2,4段)、能固定目标种的观察段组成(图3中3段)。水流由水泵输入,流速由电脑控制的阀门调节,水流由末端3根直立水管流出(图3中5段)。该装置能够将鱼类身体固定,并沿鱼身纵向产生单向流。为避免手术后鱼类创口感染,将水槽水位控制在淡水鳗鱼头部创口下方,水深约5cm。淡水鳗鱼固定在图3中标号为3的部分,通过调节水流来观察神经纤维的反应。水流由0.15 cm/s随机调节到0.5cm/s,1 cm/s,1.5cm/s,2 cm/s,2.5 cm/s,3 cm/s,3.5cm/s,4cm/s,以观察神经纤维对流速的刺激反应。研究人员对95条神经纤维进行了测试,结果有70条对流速变化有反应,25条没有明显反应。Voigt等[7]认为存在着对水流敏感的神经纤维和不敏感的神经纤维,结合已有的研究成果,得出结论:对流速敏感的神经纤维属于体表神经丘;对流速不敏感的神经纤维属于侧线管神经丘。这就验证了神经丘能够感觉流速的变化,同时说明神经丘存在分工。

图3 Voigt试验流场产生装置Fig.3 Experimental arrangement for generating flow field in Voigt's experiment
实验室的研究说明了鱼类感觉水流的生理机理,而在鱼类栖息地和水工建筑物附近,研究人员发现鱼类对存在特定水流条件的区域有着偏好,在这些特定水流条件区域内速度梯度可以作为研究鱼类行为响应的参数。Crowder等[9]总结了Fausch等[10-11]的研究成果后发现,在河鳟的栖息地中速度梯度是重要的水流特征;Smith等[12]发现速度梯度是幼鲑栖息地的重要特征;Haro等[13-14]认为洄游鱼类能够感觉在溢洪道拦鱼栅附近的速度变化,并指出在该区域速度梯度可以作为研究鱼类感觉的特征量。
2.2 鱼类对流速的偏好
鱼类对于流速响应行为的研究主要是鱼类对流速的偏好研究,即趋流性研究,以及趋流性研究具体指标——感应流速、偏好流速和极限流速的测量。鱼类的趋流性是指鱼类能根据水流的方向和速度随时调整自身的游向和游速,使自身保持逆流游泳状态或长时间停留在某一特定的位置。
感应流速是指鱼类刚刚可以产生反应的流速值,又称为起点流速。偏好流速是指在鱼类能克服的流速范围内对它们游动最为适合的流速范围,鱼常喜欢聚集在这个流速范围内顶流前进。极限流速是指鱼类所能克服的最大流速。
趋流性的研究方法主要是鱼类游泳速度的测定,相关试验多采用Brett[15-16]提出的2种测定方法:恒定流速法和流速递增法。侯轶群[17]用流速递增法对长薄鳅、中华沙鳅等进行了极限流速的测试,试验在平坡水槽中进行,水流由水泵输入,通过控制水槽尾门开度调节水槽内的流速和水位,试验装置见图4。通过试验测定长薄鳅的极限流速为0.6m/s,而中华沙鳅的极限流速大于0.6m/s。此外,张硕等[18]利用循环回流水槽研究了黑鮶幼鱼的趋流性,结果表明,黑鮶幼鱼的感应流速为0.10~0.15 m/s,偏好流速为0.20~0.45m/s,极限流速为0.6~0.7m/s,认为偏好流速和极限流速与鱼类体长的关系密切。赵希坤等[19]对鲤、鲫、鲢、草鱼、梭鱼、团头鲂 鮊、乌鳢、鲶等鱼类进行了类似的水槽试验,用以研究不同鱼类的趋流性,试验结果表明,试验鱼类的感应流速在0.2m/s左右,偏好流速在0.3~0.8m/s之间,不同种类鱼类的极限流速差别较大。鱼类表现出不同的趋流性可能与鱼类栖息地水流环境有关,栖息在河流中的鱼类其极限流速往往大于栖息在湖泊中的鱼类。
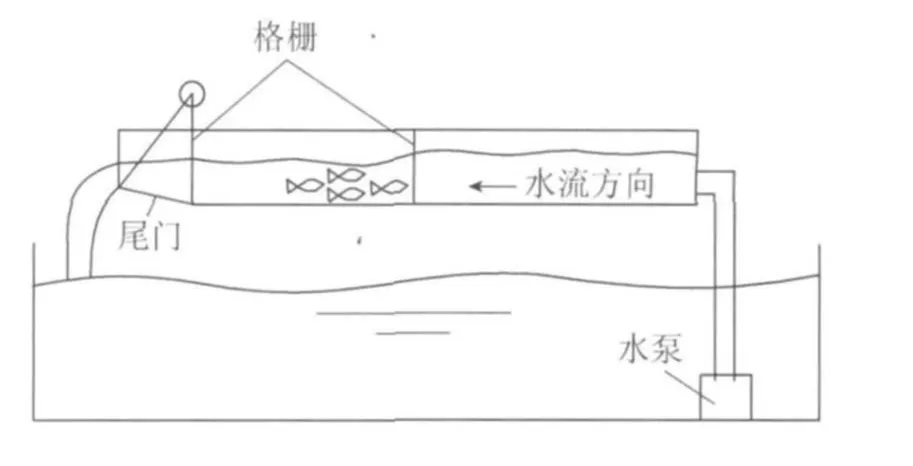
图4 鱼类游泳能力测试装置Fig.4 Experimental arrangement for testing swimming capacity of fish
3 鱼类对压力变化的响应
侧线系统的体表神经丘作为鱼类对于水流速度和强度的感受器,还能够感觉压力变化。Coombs等[20]对此进行了研究:以震动偶极子作为压力的发生源,探求鱼类侧线管神经丘对压力梯度的感觉模式;以金鱼为试验对象,假设2个侧线孔之间只有1个神经丘,侧线管神经丘的反应与外部的压力梯度成比例,以此为基础对沿金鱼侧线孔的压力空间分布进行了数值模拟,并将数值模拟结果与试验水槽中实测震动偶极子沿侧线压力分布相比较。试验方法为将震动的偶极子沿金鱼身体纵向移动,比较产生的压力梯度振幅与金鱼神经纤维的反应。对鱼类处理时,先进行麻醉,再把金鱼固定,在口中插入水管以供给水源,并在其头部开孔,露出侧线神经纤维末梢,将末梢与微电极管相连。然后移动震动源,金鱼的神经纤维会作出反应,如图5所示。神经纤维的反应由微电极管传出,经过放大就可以用作试验分析。在比较了震动偶极子产生的压力梯度振幅与微电极管传出的讯号后,认为金鱼对于身边震动产生的压力梯度有反应,感觉压力梯度的是侧线管神经丘。
在鱼类栖息地环境中,水流运动或昆虫产生震动十分常见,鱼类通过感觉震动源来选择游泳路径或觅食,因此有必要研究鱼类对震动源的感觉机理。Engelmann等[6]通过循环水槽试验来研究鱼类对于震动源的反应,以鲋鱼和虹鳟鱼为试验对象。首先实施手术处理,在头部侧线神经进入骨髓处开孔,露出侧线神经,并与装有KCl溶液的玻璃微细管相连,用以获取侧线神经反应。然后在鱼背上用胶粘上1个树脂玻璃棒以控制试验鱼的位置,在鱼尾系绳,防止试验鱼侧向移动。试验鱼置于水槽时,通过固定装置向鱼嘴注入水流,以保持鱼类正常呼吸。循环水槽水流为单向流,水流由马达带动螺旋桨产生,流速为10cm/s。采用PIV技术,安置激光发射器以及摄像机来测量流速。震动源的振动频率为50Hz,放置在距鱼类身体表面6~8mm的位置,试验设备布置如图6所示。经过试验,认为存在2种侧线神经纤维,一种是支配体表神经丘的侧线神经纤维,另一种是支配侧线管神经丘的侧线神经纤维。在单向水流中支配体表神经丘的神经纤维对水流有感觉,对震动没有反应;支配侧线管神经丘的神经纤维对水流没有明显反应,但对震动有反应。因此,得出结论,体表神经丘感觉震动的能力在流动的水流中会减弱,而侧线管神经丘在静止与流动的水流中都能对震动产生感觉。

图5 震动源移动方向以及震动源相对于鱼类位置示意图Fig.5 Schematic diagram of vibration source motion and position of vibration source relative to fish
4 鱼类对紊动和切应力的响应行为试验
4.1 紊动对鱼类的影响
紊动尺度及其作用范围影响着鱼类的行为。小尺寸的紊动会造成鱼类局部身体伤害;而大尺寸紊动则会带动鱼类起伏旋转。如果鱼类在大尺度紊动的水流内停留时间过长,就会迷失方向、丧失平衡,最终导致其游泳能力降低。王得祥[21]以鲫鱼为研究对象,利用震动格栅产生紊流,分析鱼类受到紊动产生的影响。经过试验,发现适合鲫鱼生存的紊动强度为0.56~6.64cm/s,这时鲫鱼能够在水中保持正常的游泳姿态;将紊动强度提高到7.04cm/s以上,鲫鱼不能保持正常游泳姿态;紊动强度大于9.09cm/s时鲫鱼产生了明显的伤害,表现为鱼眼受损、鱼鳞脱落、黏膜破坏等;进一步提高紊动强度到12.48 cm/s以上,水流中会出现大尺度漩涡,漩涡把鲫鱼包裹在内,使鲫鱼随着漩涡不停旋转,随后导致鲫鱼死亡。
天然河道中一般情况下不会有较大的切应力使鱼类受伤,但是在水轮机中,特别是边界层等处,流速梯度大,鱼类会受到伤害。具体来说,在鱼类栖息的天然河道中,紊动尺度可以小于鱼类的体长,如鲑鱼游动时带动水流而产生的紊动;也可以远大于鱼类体长,如大范围的水流冲击。对于洄游鱼类,Coutant等[22]认为在河流中经过长时间的进化,洄游鱼类能够利用天然河流中的紊动来减少洄游过程中的代谢能量消耗。如果这种假设是正确的,那么对于洄游中的鲑鱼一定存在最适宜的紊动条件。如果紊动过大(尾水渠、尾水管或泄洪道中),鱼类会受伤或失去方向感;如果紊动太小(如水库蓄满时),鱼类难以感知洄游方向,无法利用大范围低强度的紊动来减少自身能量消耗。对于非洄游鱼类,一些俄国学者研究了它们在低流速小强度的紊动中保持自身位置的能力。如Pavlov等[23]发现提高紊动强度会降低鱼类的极限流速,紊动较大的水流带有更多的动能,因此认为鱼类在有紊动的水流中要保持自身位置不变,就要克服紊动的影响,这意味着消耗更多的能量。然而Pavlov等[24]在后期的研究中发现处于饥饿状态的鱼类更喜欢紊动的水流,这可能是由于紊动的水流中有更多的捕食机会。由于种类和生活习性的不同,非巡游鱼类对紊流程度的偏好大相径庭。一般来说,栖息在河流中的鱼类偏好紊动程度较大的水流,而栖息在湖泊中的鱼类偏好紊动强度小的水流,这与鱼类神经丘分布不同是一致的。如前所述,由于神经丘的功能不同,栖息在河流中的鱼类有发达的侧线管道;栖息在湖泊中的鱼类体表神经丘分布广,侧线管道不发达。

图6 Engelmann试验设备Fig.6 Engelmann's experiment arrangement
然而在水轮机中,水力学条件复杂,紊动强度大,不同尺度的紊动对鱼类造成的伤害不同。在水轮机通道中小尺度的紊动无处不在,这种广泛存在的小尺度紊动会扭曲和挤压鱼类,造成鱼类身体局部受伤。大尺度的紊动在尾水渠和尾水管中最为明显,在这些地方水流横截面扩张,流速降低,并且受到水轮机转轮带动水流的冲击作用,流态十分复杂。鱼类通过这些区域时难以感应如此复杂的水流条件并作出反应,因而会迷失方向。研究表明,紊动导致的方向丧失或许对鱼类没有直接伤害,但迷失方向的鱼类进入尾水渠后易于被天敌捕食。
4.2 切应力对鱼类的影响
天然河道中的切应力一般不大,不会对鱼类造成严重伤害;水轮机、尾水渠和泄洪道等处,水流条件复杂,紊动强度大,产生的切应力往往对通过其中的鱼类造成伤害,甚至致死。然而鱼类也会表现出对特定切应力区域的偏好,Faler等[25]发现某些狗鱼(North pikeminnow)会聚集在水轮机出口或者鱼道入口等切应力较大的区域。
在水轮机中,切应力作用明显的区域为水流与固体交接边界,如转叶附近,这里水流大幅度减速,由此产生较大的切应力。由于速度急剧变化而产生的切应力有着相当大的内能,这些能量一部分随着紊流漩涡消散。相对于切应力,紊动对鱼类造成的伤害可以忽略。有研究表明,鱼类身体的不同部分处在不同的流速区域,不同的流速区域之间流速梯度很大,这就产生了相当大的切应力,因而鱼类身体会被扭曲造成伤害。
5 鱼类对综合水动力特征的响应——SVP假设
由于鱼类栖息地水流条件的复杂性,现有的研究多集中在鱼类对于单一水动力特征的响应,无法揭示鱼类在自然条件下选择游泳路径的机理,Goodwin等[26]针对这方面的空白提出了SVP假设,并将其应用于检验鱼道的过鱼效率,结果表明该假设能够反映出鱼类对于游泳路径的选择规律。
5.1 定义总水力应变的概念
SVP假设定义了总水力应变。如前所述,在鱼类的栖息地中,紊动和漩涡是极其常见的,鱼类能够感觉这种常见的水流运动并作出响应。但由于紊动是随机过程,难以定量描述,这就给研究工作带来困难。为解决这个难题,Goodwin等[26]在SVP假设中采用了1个新的标量——总水力应变(total hydraulic strain)。总水力应变以描述流场运动的欧拉法为基础,表示为速度梯度张量行列式中各项绝对值的和,即三维坐标系中速度在各方向上梯度的绝对值相加,得到标量S1。由于速度梯度张量矩阵可表示为正应变张量、角速度张量和角变形张量之和,因而称S1为总水力应变。它反映了流场区域内控制体变形情况,包含了紊动的水力特征,而且能够在CFD(computational fluid dynamic)模型中计算。

式中:S1——总水力应变;u,v,w——x,y和z方向的速度分量;εxx,εyy,εzz——x,y,z方向的正应变率;εyz,εzy— —x平面的切应变率,εzy=εyz;εxz,εzx— —y平面的切应变率,εxz=εzx;εxy,εyx——z平面的切应变率,εxy=εyx;ωxy,ωyz,ωxz,ωyx,ωzy,ωzx— —绕x,y,z轴旋转的角速度,且 ωxy=-ωyx,ωyz=-ωzy,ωxz=-ωzx。
式(1),(2),(3)分别为为正应变张量、角速度张量和角变形张量。式(4)为由正应变张量、角速度张量和角变形张量相加得到的速度梯度张量。式(5)为总水力应变,表示为速度梯度张量行列式中各项绝对值的和。
5.2 栖息地流态的特征
SVP假设在鱼类栖息地水流中可归纳为2种流态:(a)在固体边界附近产生的边界层流(wall-bound flow);(b)在河道中水流受到障碍物(如突出的岩石或树桩)产生的自由切应力流(free-shear flow)。这2种流态中,总水力应变和速度的变化表现出不同的特征。在边界层流中,沿着边界的法向方向总水力应变增加,速度值(velocity magnitude)减少;在自由切应力流中,沿着上游来流方向总水力应变和速度值都增加。结合鱼类机械感觉系统中2种神经丘的不同功用,侧线管神经丘能够感觉速度梯度变化;体表神经丘能够感觉流速的强度和方向;鱼类的机械感觉系统能够觉察总水力应变和速度值的变化,从而“辨认”出这2种流态。此外SVP假设还考虑:(a)鱼类竖直方向的运动,鱼类调整鱼鳔的体积来实现上浮或下潜。(b)鱼鳔体积的变化与静水压力(hydrostatic pressure)相关。在水体表面附近静水压力小,鱼类能够吸气扩大鱼鳔体积;在较深处静水压力大,鱼类不能获得空气,只能通过鱼鳔的膜过滤血液中的气体,因此将静水压力作为假设中的特征量。
5.3 鱼类接受水动力刺激方式
对于鱼类接受水流刺激的方式,SVP假设采用了Weber-Fechner定律,该定律认为感觉的差别阀限随原

式中:φ——原刺激量;Δφ——当前的差别阀限,也称最小可觉差(just noticeable difference);C——Weber率,代表特定感觉道的定值。
在研究动物的判别能力上,Weber定律认为只有在当前刺激强度与动物已经适应的刺激强度之差超过了最小可觉差时,动物才能够感觉到刺激强度的变化。在SVP假设中,Goodwin等[27-28]将Weber定律应用于研究鱼类对水力特征量的感受方式,提出在当前特征量强度与鱼类已经适应了的刺激强度存在差别是鱼类感觉到刺激的前提,但只有当这个差别超过了阀限鱼类才能够觉察特征量的变化。
5.4 鱼类对水动力刺激的响应
在SVP假设中,Goodwin等将k,d定义为当前刺激强度与鱼类已经适应刺激强度之差;把k1与k2分别定义为鱼类能够察觉的边界层流总水力应变和自由切应力流总水力应变的阀限,并且认为k1小于k2;定义k3为与鱼类浮潜深度变化阀限。经研究发现鱼类游泳路径选择规律:
a.在没有障碍物的直道水槽、水流为均匀流状态下,觉察到与边界层流相关的总水力应变增加时(k1<k<k2),朝着水流速度增加的方向游动。Coutant等[29]的研究支持了这个发现,他们注意到洄游鱼类接近大坝时总是偏好流速较大的区域。洄游鱼类通过这种行为减少洄游时间,进而降低能量消耗。
b.河道中有障碍物影响水流输送并且导致紊动产生的情况下,鱼类觉察到与自由切应力流相关的总水力应变增加时(k1<k2<k),鱼类朝着水流速度减小的方向游动,或者逆水游泳,这样可以使鱼类避免在总水力应变较大区域中的停留时间过长,从而减少被高强度紊流伤害的几率。
c.当鱼类感受到水深变化(即静水压强)增加时(k3<d)时,鱼类游向前期已适应的深度。
Goodwin等将总水力应变(total hydraulic strain)、速度值(velocity magnitude)和静水压力(hydrostatic pressure)这3个水力特征量结合在一起,根据Weber定律制定了鱼类选择路径的规则,并将其应用于检验鱼道的过鱼效率,结果表明该假设能够反映鱼类对于游泳路径的选择[28,30]。来刺激量的变化而变化,且表现为一定的规律性,表示为
6 结 语
鱼类感受器作用机理的研究已经比较成熟,如Baker研究鱼类神经丘分工的试验,Coombs的关于鱼类感觉压力梯度的试验,Engelmann对于鱼类感觉震动源的试验,这些研究从鱼类学角度揭示了鱼类感受水动力刺激的生理机理。对于定义影响鱼类行为的水动力特性,现有的研究主要从流速方面来揭示鱼类的趋流性机理,但是在鱼类的栖息地中,水流条件十分复杂,在实验室中研究趋流性所构造的均匀流并不常见,这就要求提出能够综合反应水流特征的参数对栖息地水流进行整体的描述,如Goodwin提出的总水力应变的概念,将速度梯度绝对值的和作为水动力指标,从而避免了对紊动的讨论。在定义水动力指标时,要将鱼类能够直接感受到的水动力特征,如速度、压力以及速度梯度等,与能够反应水流整体特征的水动力特征量联系起来,同时避免对水流随机过程如紊动或漩涡的讨论,这样才能得到可以量化的指标,用于分析计算,这是鱼类学与水力学的结合点[31],也是研究鱼类对水动力环境特征响应的发展方向。
[1]KANTERM J,COOMBS S.Rheotaxis and prey detection in uniform currents by Lake Michigan mottled sculpin(Cottus bairdi)[J].Journal of Experimental Biology,2003,206(1):59.
[2]MARUSKA KP.Morphology of the mechanosensory lateral line system in elasmobranch fishes:ecological and behavioral considerations[J].Environmental Biology of Fishes,2001,60(1):47-75.
[3]HUDSPETH A J.Transduction and tuning by vertebrate hair cells[J].Trends in Neurosciences,1983,6:366-369.
[4]BLECKMANN H.3-D-orientationwith the octavolateralis system[J].Journal of Physiology-Paris,2004,98(1-3):53-65.
[5]FLOCKM BK,WERSGLL J.A study of the orientation of the sensory hairs of the receptor cells in the lateral line organ of fish,with special reference to thefunction of the receptors[J].The Journal of Cell Biology,1962,15(1):19.
[6]ENGELMANNJ,HANKE W,BLECKMANNH.Lateral line receptionin still-and runningwater[J].Journal of Comparative Physiology A:Neuroethology,Sensory,Neural,and Behavioral Physiology,2002,188(7):513-526.
[7]VOIGT R,CARTON A G,MONTGOMERY J C.Responses of anterior lateral line afferent neurones to water flow[J].The Journal of Experimental Biology,2000,203(Pt 16):2495.
[8]CARTON A G,MONTGOMERY J C.Responses of lateral line receptors to water flow in the Antarctic notothenioid,Trematomus Bernacchii[J].Polar Biology,2002,25(10):789-793.
[9]CROWDERD W,DIPLASP.Vorticity and circulation:spatial metrics for evaluating flow complexity in stream habitats[J].Canadian Journal of Fisheries and Aquatic Sciences,2002,59(4):633-645.
[10]FAUSCH KD.Experimental analysis of microhabitat selection by juvenile steelhead(Oncorhynchus mykiss)and coho salmon(O.kisutch)in a British Columbia stream[J].Canadian Journal of Fisheries and Aquatic Sciences,1993,50(6):1198-1207.
[11]HAYES JW,JOWETTIG.Microhabitat models of largedrift-feedingbrowntrout in three New Zealand rivers[J].North American Journal of Fisheries Management,1994,14(4):710-725.
[12]SMITH D L.The shear flow environment of juvenile salmonids[D].Moscow:University of Idaho,2003.
[13]HAROA,ODEH M,NOREIKA J,et al.Effect of water acceleration on downstream migratory behavior and passage of Atlantic salmon smolts and juvenile American shad at surface bypasses[J].Transactions of the American Fisheries Society,1998,127(1):118-127.
[14]FLETCHER RI.Flows and fish behavior:largedouble-entry screeningsystems[J].Transactionsof the American Fisheries Society,1994,123(6):866-885.
[15]BRETT J R.The respiratory metabolism and swimming performance of young sockeye salmon[J].Fish Res Board Can,1964,21:1183-1226.
[16]BRETT J R.Swimming performanceof sockeye salmon(Oncorhynchus nerka)in relation to fatigue time and temperature[J].Fish Res Board Can,1967,24:1731-1741.
[17]侯轶群.洄游鱼类的过坝能力试验与鱼道数值模拟[D].南京:河海大学,2010.
[18]张硕,陈勇.黑鮊幼鱼趋流性的初步研究[J].上海水产大学学报,2005,14(3):282-287.(ZHANG Shuo,CHEN Yong.Preliminary study on the rheotaxisof juvenileSebastodes fuscescens[J].Journal of Shanghai Fisheries University,2005,14(3):282-287.(in Chinese))
[19]赵希坤,韩桢锷.鱼类克服流速能力的试验[J].水产学报,1980,4(1):31-37.(ZHAO Xikun,HAN Zhene.Experiments on the current overcoming ability of some freshwater fishes[J].Journal of Fisheries of China,1980,4(1):31-37.(in Chinese))
[20]COOMBSS,HASTINGSM,FINNERANJ.Modeling and measuring lateral line excitation patterns to changingdipolesourcelocations[J].Journal of Comparative Physiology A:Neuroethology,Sensory,Neural,and Behavioral Physiology,1996,178(3):359-371.
[21]王得祥.水流紊动对鱼类影响试验研究[D].南京:河海大学,2007.
[22]COUTANT C C,WHITNEY R R.Fish behavior in relation to passage through hydropower turbines:a review[J].Transactions of the American Fisheries Society,2000,129(2):351-380.
[23]PAVLOV D S,SKOROBOGATOV M A,SHTAF L G.Influence of degree of stream turbulence on the magnitude of the critical current velocity for fish[J].Doklady Biological Sciences,1982,267:560-562.
[24]PAVLOV D S,LUPANDINA I,DEGTYAREVA N G,et al.Role of turbulence in the distribution of downstream migrating young fishes(early larval stages)inwide and narrow channels[R].New York:Consultants Bureau,1995,341:211-215.
[25]FALERM P,MILLER LM,WELKEKI.Effects of variation in flow on distributions of northern squawfish in the Columbia River below McNary Dam[J].North American Journal of Fisheries Management,1988,8(1):30-35.
[26]NESTLER JM,GOODWIN RA,SMITH D L,et al.Optimum fish passage and guidance designs arebased in thehydrogeomorphology of natural rivers[J].River Research and Applications,2008,24(2):148-168.
[27]GOODWIN R A,NESTLER JM,ANDERSON J J,et al.Forecasting 3-D fish movement behavior using a Eulerian-Lagrangian-Agent method(ELAM)[J].Ecological Modelling,2006,192(1-2):197-223.
[28]GOODWIN R A,ANDERSON J J,NESTLER JM.Decoding 3-D movement patterns of fish in response to hydrodynamics and water quality for forecast simulation[M].Singapore:World Scientific Publishing Company,2004.
[29]COUTANTC C,WHITNEY R R.Fish behavior in relation to passage through hydropower turbines:a review[J].Transactions of the American Fisheries Society,2000,129(2):351-380.
[30]WEBER L J,GOODWIN RA,LIS,et al.Application of an Eulerian-Lagrangian-Agent method(ELAM)to ranKalternative designs of a juvenile fish passage facility[J].Journal of Hydroinformatics,2006,8(4):271-295.
[31]杨宇,严忠民,乔晔.河流鱼类栖息地水力学条件表征与评述[J].河海大学学报:自然科学版,2007,35(2):125-130.(YANG Yu,YANZhongmin,QIAOHua.Description and review of hydraulic conditions of fish habitats[J].Journal of Hohai University:Natural Sciences,2007,35(2):125-130.(in Chinese))
猜你喜欢
科学大众(2020年12期)2020-08-13 03:22:32
水电站设计(2018年1期)2018-04-12 05:31:58
中国公路(2017年18期)2018-01-23 03:00:38
水利规划与设计(2016年9期)2017-01-15 14:00:51
铁道通信信号(2016年7期)2016-06-06 02:21:02
小学阅读指南·低年级版(2016年8期)2016-05-14 13:24:45
电网与清洁能源(2015年5期)2015-12-29 11:52:57
中国石油大学学报(自然科学版)(2015年2期)2015-11-10 06:08:26
西安建筑科技大学学报(自然科学版)(2014年4期)2014-11-12 05:15:24
城市道桥与防洪(2014年7期)2014-02-27 07:28:03
