
向心训练与离心训练对大鼠腓肠肌H反射的影响
2012-03-06 04:56周思红
中国体育科技 2012年2期
周思红,刘 宇
1 前言
不同训练方式能够对神经肌肉系统产生不同的影响,这其中,离心训练对肌肉影响的研究相对较多,而对神经反射兴奋性的相关报道则较少,特别是H反射方面的影响更少[2,7,14,16,28]。
Ghristian[20]研究人体从不同高度进行落地反跳时,其比目鱼肌H反射的变化,发现相同高度下H反射在触地阶段要显著小于下落阶段,且H反射随着下落高度的增加而逐渐减小,显示人体的这种反射调节是为了调整肌腱刚度,以便肌腱在保持较大刚度的同时,避免由于超载负荷而可能造成的损伤。田石榴[12]研究发现,8周负重超等长训练后,H反射中的主要指标Hmax/Mmax值降低,显示负重超等长使α运动神经元终池兴奋性降低。
诚然,从20个世纪开始就有不少学者对H反射进行了研究[17,21,23,29,30],但研究涉及运动训练方面,特别是离心训练对神经肌肉系统的影响,其机制到目前为止尚不清楚。因此,本研究的目的,为了解神经系统在不同运动方式后的适应情况,通过探究向心与离心训练8周后大鼠H反射特征的变化,了解不同训练方式对神经肌肉系统的影响。
2 研究对象和方法
2.1 研究对象
雄性SD健康大鼠72只,平均体重(180~200)g,购自上海西普尔-必凯实验动物有限公司。大鼠在实验室适应3天后被随机分为安静对照组(C组)向心训练组(EC组)和离心训练组(EE组)(所有分组n=24)。各组大鼠经独立样本检验,体重无显著性差异。大鼠分笼饲养,每笼不超过5只,对每组大鼠采取身体不同部位涂抹3%~5%的苦味酸来进行标识,涂后毛发呈黄色。训练房的室温保持在22℃左右,相对湿度保持在50%~60%,昼夜节律用日光灯控制,每日光照时间为8:00AM~8:00PM。训练期间,训练房一直保持通风、干净、整洁。大鼠自由饮食,每周2次定时对其称重并记录[3,8,9]。
2.2 研究方法
2.2.1 向心和离心训练动物模型的建立
对照组和运动组处于相同条件的房间内,但不进行任何形式的运动,每天正常饮食。正式实验前经预实验测定运动后即刻大鼠尾部的血乳酸值,评定在不同坡度跑台上大鼠的运动强度,然后确定运动负荷和运动方案,争取使EE组大鼠和EC组大鼠运动负荷相似。EC组和EE组大鼠在进实验室第3、4天,每天进行1次速度为5~10m/min,时间为5~10min,坡度为0°的跑台运动进行适应性训练,第5、6天使EC组大鼠每天进行1次速度为10~15m/min,时间为10~15min,坡度为4°的跑步训练。第5、6天使EE组大鼠每天进行1次速度为10~15m/min,时间为10~15min,坡度为-14°的跑步训练[10,15,18]。使两组大鼠熟悉在动物跑台上进行上坡跑和下坡跑训练。第7天两组休息。正式实验在第2周开始,到第9周结束,均在下午2点开始训练,训练总时间持续8周,其中训练组大鼠周一到周五训练5天,周六和周日休息。
2.2.2 训练方案
训练组大鼠的训练方案设计为,EC组大鼠从静止开始,在坡度为4°的跑台上,每天进行1次速度为19~23m/min、训练时间为25~40min的向心运动。EE组大鼠从静止开始,在坡度为-14°的跑台上,每天1次速度为17~21m/min、总时间25~40min的离心运动(表1,表2)。
2.2.3 取材方法
取材前一天,对取材环境进行全面消毒,并保证取材环境的卫生整洁、干净,温度和适度保持正常。在最后一次运动结束后24h取材,3组同日取材。大鼠处死前12h禁止饮食,称重,按400mg/kg体重剂量,腹腔注射10%水合氯醛进行麻醉[1,4,13]。待大鼠进入深度麻醉后,取仰卧位将其固定于动物实验操作台上,常规消毒后,从大鼠内踝开始向大腿方向用剥离肌肉,在7min之内迅速钝性剥取完整的大鼠右侧腓肠肌,分离出坐骨神经,神经分离需用玻璃分针,分离过程中操作必须精细,避免使用金属器械碰到神经,防止损伤和影响标本制作。仔细制备好完整的与坐骨神经相连的腓肠肌标本。

表1 EC组具体训练安排情况一览表

表2 EE组具体训练安排情况一览表
2.2.4 H反射测试方法与步骤
首先,测大鼠右侧腓肠肌的H反射,测定H反射时,以双极银丝电极作为刺激电极,电极直接钩于坐骨神经上,统一位置选择在近坐骨神经连接腓肠肌处2~3mm处。将刺激电极通过导线连接到生物机能实验系统上的电刺激输出端,记录电极为同心针电极,将同心针电极垂直于肌纤维方向插入腓肠肌肌腹饱满处。仪器的地线通过铜线与深埋于地下的铁钉相连,使肌电图的基线受到的干扰尽量减小到最小程度。
对SD大鼠进行肌电测定时,要注意对生物机能实验系统参数的设定,设置系统采用单次电刺激诱发H波,模式为细电压,刺激波形采用方波,波宽为0.2ms,强度从0v开始,以0.005v为增量,延时100ms,通过程控进行控制。通过从小到大的电刺激强度刺激将会使H波出现,再逐渐增强刺激又将使M波出现,H波变小。然后将SD大鼠腓肠肌上采集的肌电信号记录下来[13]。通过记录腓肠肌的Mmax波和Hmax波的振幅、Hmax/Mmax之比、Mmax波出现的最大上升速度和最大下降速度、Hmax波出现的最大上升速度和最大下降速度等指标来反映H反射的有关情况[19,22,24,27]。
2.3 数据统计方法
3 实验结果
3.1 3组大鼠腓肠肌H反射测试图

图1 EC组肌电图

图2 EE组肌电图

图3 C组肌电图
经过对3组大鼠腓肠肌的肌电测试,可以对H反射的主要指标数据进行采集,3组大鼠测试的肌电图如图1~图3所示。
3.2 3组大鼠腓肠肌H反射各指标测试结果
由表3和图4~图6所示,通过对H反射的比较发现,Mmax波显示EE组显著大于C组和EC组,但EC组与C组之间无显著性差异。Hmax波显示EE组显著大于C组。Hmax/Mmax值显示3组之间均无显著性差异。从Mmax波与Hmax波出现的时间发现,EE组Mmax波与Hmax波出现时间均要显著迟于EE组大鼠,但与C组之间无差异,同时EC组与C组之间也无显著性差异。

表3 H反射测试结果一览表

图4 M波和H波最大振幅示意图
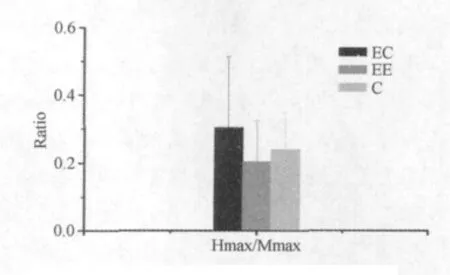
图5 H波和M波最大振幅比值示意图
4 分析讨论
目前,检测神经系统中周围神经功能的客观指标的方法之一是采取肌电图检查,这种检测手段已被许多临床医生以及广大科研人员所接受[6,11]。其中在肌电图检查中,H反射是检查中的一个重要指标,它是通过对支配腓肠肌的坐骨神经采用脉冲电流或机械刺激引起腓肠肌兴奋,然后对采集的电信号进行记录。通过检查坐骨神经的末梢运动神经纤维传导速度、运动终板的功能以及脊髓的反射功能等指标,来反映周围神经的功能。其经常使用的指标有潜伏期、Mmax、Hmax、Hmax/Mmax值等。通过对大鼠进行H反射检测,研究运动方式对神经系统的影响相对较少。本组实验旨在进行这方面的探索,以期能为进一步检测奠定基础。

图6 M波和H波最大振幅出现时间示意图
4.1 向心与离心训练对Mmax波和Hmax波的影响
从Mmax波的测定结果发现,C组与EE组之间有特别显著性差异,EC组与EE组之间也有显著性差异,C组与EC组之间无显著性差异。从Hmax波的测定结果发现,C组与EE组之间有显著性差异,而C组与EC组之间,EC组与EE组之间均无显著性差异。Hmax/Mmax值测定结果表明,3组之间无显著性差异。研究表明,M波是通过电刺激坐骨神经直接兴奋α运动神经元轴突而引起的H反射波,M波的大小与神经-肌肉接点以及肌纤维的功能状态有密切关系。
本研究选择的多道生理记录仪设定的刺激参数条件合适,经记录的数据能准确采集到大鼠坐骨神经-腓肠肌检测H反射时的Mmax波和Hmax波,测试的数据结果精确,读取方便,可重复性比较强,误差极小。有关数据结果呈正态分布。因为测定的H波是由电刺激Ia类传入神经纤维产生的反射波。因此H波的振幅大小与神经支内Ia类传入神经纤维的含量多少有关。运动单位中的运动神经元多少直接影响到骨骼肌电位的高低程度,电压高低与它兴奋肌纤维的密度有直接关系,电压越高,兴奋的肌纤维数就越多,密度也就越大。本研究通过测定大鼠腓肠肌H反射的Hmax波,可验证大鼠周围神经终支内Ia类传入神经纤维的含量。
因为实验在相同的刺激条件下,不同组别大鼠腓肠肌的Hmax波的平均值大小,反映了该组大鼠被兴奋的Ia类传入神经纤维电活动的大小程度,Hmax波平均振幅之间的差异,能够反映出3组大鼠神经支之间Ia类传入纤维含量多少的差异。分析认为,只有电刺激较多的Ia类传入神经纤维,才能导致兴奋更多的运动神经元,使Hmax波有较高的幅值[25]。影响H反射的因素较多,主要分为内部因素和外部因素。而内部诸因素中,最重要的影响因素是引起神经反射的兴奋阈值以及被兴奋的神经纤维数量多少。因为H反射属于低阈值反射,因此,当脉冲电刺激强度比较弱时,记录电极首先会记录到H波,随着刺激强度逐渐增加,H波也会增大,但刺激强度过高超过一定范围以后,H波反而会随着刺激强度的增加,逐渐下降。只有当刺激强度增加到足以引起阈值较高的α运动神经元兴奋时,则会产生M波,如果刺激强度继续增加,将会出现H波消失,M波达到最高。在实验过程中可以通过这个特点来确定H反射。在相同的刺激条件下,如果Ia类传入神经纤维的数量越多,将会使得兴奋α神经元的数量也就越多,导致H波振幅也越高,因此,从本实验结果发现,因为EE组的Hmax波显著高于C组,也反映出经过离心训练可能导致EE组SD大鼠体内坐骨神经干Ia类传入神经纤维的数量出现增多的情况。
4.2 向心与离心训练对Hmax/Mmax的影响
实验中测定的Hmax/Mmax表示经多道生理记录仪测定的腓肠肌Hmax波与刺激坐骨神经产生的Mmax波的比值。研究报道,运动训练中的Hmax/Mmax大小变化,除受脊髓的运动神经元兴奋性影响外,还受Ia类传入纤维的突触前抑制(presynaptic inhibition-PSI)的影响。离心训练因为骨骼肌在收缩状态下被拉长,将会导致本体感受器的传入增加,支配骨骼肌活性的Ia运动神经元的氧化酶活性增加等。这些因素都可以使大鼠离心训练后脊髓运动神经元的兴奋性增加[20]。研究认为H反射可以反映α运动神经元群的兴奋性,无论骨骼肌疲劳后肌纤维兴奋性发生什么变化,Hmax/Mmax值也都反映α运动神经元群的兴奋性。Jayne等研究报道,人的腓肠肌在疲劳状态下Hmax/Mmax值显著性降低,H波甚至消失,研究为神经-肌肉疲劳提供了新思路。较多研究报道,M波振幅降低的原因是因为骨骼肌疲劳时肌膜的兴奋性降低所导致。就骨骼肌疲劳与H反射的关系而言,有报道认为,H反射在骨骼肌缺血疲劳的状况下振幅将会下降,如果骨骼肌持续疲劳,肌肉进行亚极量主动收缩时,H反射的振幅又会逐渐上升。Hmax/Mmax值估计受到脊髓神经元兴奋性的影响[31]。
Hmax/Mmax值降低的原因,根据动物神经生理学分析,可能因为受到电刺激后,兴奋逆向传播至脊髓α运动神经元,导致传入神经元与运动神经元进行换元的突触受到了抑制,使得肌梭的兴奋通过Ia类传入神经纤维至α运动神经元换元时出现了衰减,二次反射的兴奋性就会下降,使得H波降低,Hmax/Mmax值也相应降低。该反应有一定的生理意义[26],因为可以降低肌梭对运动神经元兴奋的负反馈调节,可以维持α运动神经元的兴奋性,从而延缓和减少神经疲劳的发生。其次,Hmax/Mmax值降低可能与机体调整平衡的能力有关。过于兴奋时抑制,过于抑制时兴奋。因为实验是在SD大鼠高级中枢麻醉状态下进行的,因此可能使脊髓的α运动神经元反应阈值产生改变,而运动训练对α神经元起到了抑制作用。分析认为,H反射属于低阈值反射,如果刺激强度已经足够大,但H波的振幅没有升高反而降低,因此可能是通过调节α运动神经元的阈值大小,来平衡兴奋和抑制过程的。Hmax/Mmax值,3组之间无显著性差异,表明3组大鼠测定的Hmax波与Mmax波之间呈现同时增大或缩小情况。
4.3 向心与离心训练对Mmax波和Hmax波出现时间的影响
从Mmax波和Hmax波出现的时间测定结果发现,EC组与EE组之间出现显著性差异,C组与EC组之间无显著性差异。表明EE组Mmax波和Hmax波出现都要显著晚于EC组。从研究结果分析,估计这种情况出现的原因是由于运动训练导致大鼠的H波和M波发生改变,特别是离心训练后由于腓肠肌的肌力增强,致使Hmax波出现的时间推迟。另外,Mmax波出现推迟,推测与α运动神经元的抗电刺激能力提高有一定关系。
测定发现,SD大鼠的后肢H反射具有与人类相似的波型特征,但因为大鼠后肢比较短,致使刺激坐骨神经的电极与腓肠肌上的记录电极距离较近,造成刺激波与M波的间距较小,潜伏时间也较短的特点。而H波也有一定的特点,随着脉冲电刺激的逐渐增强,它也会逐渐增加振幅,随后当H反射的振幅达到最大,如果继续增大脉冲电刺激时,就会出现M波,当M波增大时,H波则逐渐降低,强度达到一定程度时,H反射消失。根据不同实验的研究目的与设计要求,进行诱发SD大鼠腓肠肌肌电图时采用的方法也不同。经过研究人员的努力,目前对H反射的研究和认识已比较深入,因为H反射作为一种电刺激诱发的脊髓的单突触牵张反射,可以帮助人们判断脊髓中α运动神经元的兴奋性高低,所以,根据H反射的变化情况可作为判断脊髓的相应节段,特别是可作为评判此相应节段α运动神经元功能状态的一种比较客观的指标。
5 结论与建议
训练后大鼠神经肌肉系统兴奋性增加,特别是离心训练使Mmax波明显增加,提示离心训练对改善肌肉组织的兴奋性有一定作用;离心训练后,Hmax波增加,说明离心训练对改善Ia类传入神经纤维的功能有一定作用;Hmax波和Mmax波出现的时间推迟,乃α运动神经元的兴奋性阈值增高所致,提示离心训练后,α运动神经元的抗电刺激能力提高。因此,离心训练可以使大鼠的神经肌肉系统适应性得到改善。
[1]崔淑芳.实验动物学[M].上海:第二军医大学出版社,2007:8.
[2]陆彩凤,王维群,李道鸿.一次力竭性离心运动对大鼠骨骼肌VEGF表达、血管增生和形态学结构的影响[J].中国运动医学杂志,2008,27(1):36-39.
[3]李世成,邵雄杰,甄达文,等.运动对肥胖大鼠血清脂联素、血脂及体脂水平的影响[J].中国运动医学杂志,2009,28(1):41-44.
[4]李俊平,徐玉明,王瑞元.常用动物运动模型的方式与发展[J].北京体育大学学报,2006,29(12):1669-1671.
[5]卢纹岱.SPSS for Windows统计分析[M].北京:电子工业出版社,2002:9.
[6]卢祖能,曾庆杏,李承晏,等.实用肌电图学[M].北京:人民卫生出版社,2000:5.
[7]马建,刘波,罗小兵,等.大强度肌肉离心训练以及抗肌肉疲劳中药对家兔体重、血红蛋白和脾脏组织结构影响的实验研究[J].中国运动医学杂志,2001,20(1):85-87.
[8]毛丽娟,许豪文.运动对大鼠肝脏GSH、GSSG含量及GSH/GSSG的影响[J].体育与科学,2004,25(1):60-63.
[9]牛燕媚,苑红,刘彦辉,等.有氧运动对胰岛素抵抗小鼠骨骼肌球形脂联素及腺苷酸活化蛋白激酶的影响[J].中国运动医学杂志,2009,28(1):36-40.
[10]邵晓梅,方剑乔,应晓明.不同针灸疗法对跑台训练大鼠海马和下丘脑内皮素受体AmRNA表达的影响[J].中国运动医学杂志,2009,28(3):307-309.
[11]尚西亮,陈世益,任惠民,等.低频电刺激过程中大鼠骨骼肌肌张力和肌电信号的变化特征[J].中国运动医学杂志,2009,28(1):29-31.
[12]田石榴.负重超等长力量训练的神经肌肉适应机制研究[D].上海体育学院博士学位论文全文数据库,2009.
[13]吴晓晴.动物实验基本操作技术手册[M].北京:人民军医出版社,2008:9.
[14]袁建琴,王瑞元,李肃反.离心运动对大鼠骨骼肌结蛋白分布和表达的影响-对骨骼肌损伤机制的研究[J].体育科学,2005,25(6):63-66.
[15]于新凯,田野.1周跑台训练对大鼠骨骼肌A-actin基因表达的影响[J].中国运动医学杂志,2001,20(1):16-18.
[16]张晓君.H反射及其临床应用的研究近况[J].临床神经病学杂志,1994,7(4):248-250.
[17]周思红.向心结合离心等速训练后的肌力变化特征[J].体育学刊,2009,16(5):104-107.
[18]ARMSTRONG R B.Eccentric exercise-induced injury to rat skeletal muscle[J].J.Appl.Physio,1983,54:80-93.
[19]ANDREAS HOLTERMANN,KARIN ROELEVELD,MORTEN ENGSTRØM,et al.Enhanced H-reflex with resistance training is related to increased rate of force development[J].Eur J Appl Physiol,2007,101:301-312.
[20]CHRISTIAN LEUKEL,ALBERT GOLLHOFER,MARTIN KELLER,et al.Phase-and task-specific modulation of soleus H-reflexes during drop-jumps and landings[J].Exp Brain Res,2008,190:71-79.
[21]EI SLIVKO,G A BOGUTSKAYA.Modulation of the soleus muscleНreflexcaused by ballistic voluntary arm movements in humans[J].Neurophysiol,2008,40(2):147-154.
[22]HEINEMEIER K M.Expression of collagen and related growth factors in rat tendon and skeletal muscle in response to specific contraction types[J].J Physiol,2007,582(3):3-16.
[23]MARC KLIMSTRA E,PAUL ZEHR.A sigmoid function is the best fit for the ascending limb of the Hoffmann reflex recruitment curve[J].Exp Brain Res,2008,186:93-105.
[24]MICHAEL J,GREY KLAUS KLINGE,CLARISSA CRONE.Post-activation depression of Soleus stretch reflexes in healthy and spastic humans[J].Exp Brain Res,2008,185:189-197.
[25]MARKUS GRUBER,ALBERT GOLLHOFER.Impact of sensorimotor training on the rate of force development and neural activation[J].Eur J Appl Physiol,2004,92:98-105.
[26]MARK MELNYK,ALBERT GOLLHOFER.Submaximal fatigue of the hamstrings impairs specific reflex components and knee stability[J].Knee Surg Sports Traumatol Arthrosc,2007,15:525-532.
[27]OLLE LAGERQUIST E,PAUL ZEHR,EVAN R L.Diurnal changes in the amplitude of the Hoffmann reflex in the human soleus but not in the flexor carpi radialis muscle[J].Exp Brain Res,2006,170:1-6.
[28]S J HEAP.Microcirculation in rat soleus muscle after eccentric exercise:the effect of nifedipine[J].Eur J Appl Physiol,2006,97:687-694.
[29]SHEILA SCHINDLER-IVENS,DAVID A,BROWN GWYN N.Soleus H-reflex excitability during pedaling post-stroke[J].Exp Brain Res,2008,188:465-474.
[30]WOLFGANG TAUBE,CHRISTIAN LEUKEL,ALBERT GOLLHOFER.Influence of enhanced visual feedback on postural control and spinal reflex modulation during stance[J].Exp Brain Res,2008,188:353-361.
猜你喜欢
体育科技文献通报(2022年3期)2022-05-23
医学综述(2021年16期)2021-12-01
阅读(科学探秘)(2019年5期)2019-07-19
教育教学论坛(2019年18期)2019-06-17
中西医结合心血管病电子杂志(2018年28期)2018-11-19
运动(2018年14期)2018-07-16
体育科学(2018年3期)2018-04-20
教育教学论坛(2018年1期)2018-01-18
中外医学研究(2017年15期)2017-06-29
心脑血管病防治(2015年3期)2016-01-15
