
橘红心大白菜形态标记的遗传及稳定性研究
2012-02-23 01:00耿新翠沈向群洪雅婷马士芳
中国蔬菜 2012年6期
耿新翠 沈向群 侯 华 洪雅婷 马士芳
(沈阳农业大学园艺学院,辽宁沈阳 110866)
形态标记指作物特定的、肉眼可见的、简单直观的外部特征,或借助简单测试即可识别的某些性状,主要包括植物学性状、农艺性状、工艺性状和抗性等。此种标记具有简单直观的特点,是遗传研究和杂种鉴定常用的传统方法(钟淮钦 等,2005)。在作物育种中利用形态标记可以提高种子纯度和选育效率(吴新杰 等,2005)。在杂种优势利用逐渐发展的今天,形态标记应用于杂种鉴别、纯度鉴定和品种权益保护具有重要意义(涂金星和傅廷栋,2001)。
球心叶为橘红色是大白菜〔Brassica campestrisL.ssp.pekinensis(Lour)Olsson〕中的稀有颜色。橘红心大白菜的球心色由1 对隐性等位基因控制,其花色与球心色的遗传关系是连锁遗传或一因多效,在转育过程中根据花色就能筛选到橘红心材料(余阳俊 等,1993;张德双 等,2003;冯辉 等,2009)。笔者在转育抗根肿病橘红心大白菜核基因雄性不育系研究过程中,通过种子低温处理后发现橘红心大白菜的子叶颜色与普通心大白菜的子叶颜色明显不同,表现为橘红色,而普通心大白菜的子叶颜色为黄色。本试验以橘红心大白菜和普通心大白菜为亲本,探讨了橘红心大白菜子叶颜色的遗传规律及其作为形态标记的稳定性,解决了橘红心大白菜转育过程中如何在早期得到橘红心性状、淘汰普通心的问题,减少了工作量。
1 材料与方法
1.1 材料
橘红心大白菜自交系(LH)由黑龙江省农业科学院提供,普通心大白菜自交系(91-12)由沈阳农业大学园艺学院提供,LH 和91-12 刚剖开后的球心色见彩版Ⅰ和彩版Ⅱ。


1.2 方法
试验于2009年2月~2010年11月在沈阳农业大学蔬菜基地进行。橘红心大白菜与普通心大白菜分别进行正反交,得到F1、F1′,F1自交得到F2,回交得到BC1F1。然后把得到的橘红心、普通心、F1、F1′、F2和BC1F1的种子分别按以下方法进行处理。
方法一:第1 组选择普通心(P1)、橘红心(P2)、F1、F1′、F2和BC1F1籽粒饱满的种子浸种催芽后,低温春化(0~3 ℃)处理,30 d 后放在阳光下2 h,统计橘红色子叶和黄色子叶,根据统计结果分析橘红心大白菜子叶颜色的遗传规律。第2 组选择普通心(P1)、橘红心(P2)、F1、F2和BC1F1籽粒饱满的种子浸种催芽后常温遮光处理,子叶展开后放在阳光下2 h,统计橘红色子叶和黄色子叶;然后分别栽到不同的穴盘里,待幼苗4~5 片真叶时移栽到大地里,10月下旬收获,调查心色;根据调查结果对橘红心大白菜子叶颜色与球心色的遗传关系进行分析。第3组选择普通心(P1)、橘红心(P2)、F1、F2和BC1F1籽粒饱满的种子浸种催芽后,低温春化(0~3 ℃)处理,30 d 后放在阳光下2 h,统计橘红色子叶和黄色子叶,然后分别栽到不同穴盘中,开花后调查花色,分析橘红心大白菜子叶颜色与花色的遗传关系。
方法二:选择普通心(P1)、橘红心(P2)、F1、F2和BC1F1籽粒饱满的种子浸种催芽后,第1 组在室温(20~25 ℃)条件下采用室内自然光和暗箱培养,子叶展开后放在阳光下2 h,调查子叶颜色。第2 组在春化条件(0~3 ℃)下采用光照(光照强度200 lx)和暗箱两种方法培养,30 d 后结束春化,放在阳光下2 h 后调查子叶颜色。
方法三:第1 组把普通心(P1)、橘红心(P2)、F1、F2和BC1F1籽粒饱满的种子浸种催芽24 h,胚根露出后在春化条件下暗箱培养;第2 组浸种催芽48 h,子叶基本展开后在春化条件下暗箱培养。30 d 后结束,放在阳光下2 h 后调查子叶颜色。
1.3 数据处理
样本容量按公式(1)计算,适合性检验按公式(2)计算。

(1)式中n为样本容量,P为目标基因发生概率;(2)式中X2为卡平方,A和a分别为显性组和隐性组的实际观察次数,n=A+a即总次数,r为理论分离比率,当α=0.05 时,X2=3.841。
2 结果与分析
2.1 橘红心大白菜子叶颜色的遗传特征
2.1.1 子叶颜色的分离规律 方法一第1 组子叶颜色调查结果显示(表1),91-12 大白菜的子叶颜色全部为黄色,橘红心大白菜子叶颜色全部为橘红色。以这两个亲本子叶颜色作为对照,正交后代(F1)子叶颜色为黄色,说明子叶的橘红色对黄色表现为隐性(见彩版Ⅲ);反交后代(F1′)子叶颜色为黄色,说明子叶橘红色的遗传不受细胞质影响。F2和BC1F1子叶颜色出现分离,经卡平方检验,F2群体中子叶黄色和橘红色的分离比为3∶1(见彩版Ⅳ);BC1F1群体中子叶黄色和橘红色的分离比为1∶1(见彩版Ⅴ)。这说明,橘红心大白菜子叶的橘红色受1 对隐性核基因控制,属于质量性状遗传。



2.1.2 子叶颜色与球心色和花色的遗传关系 从橘红心大白菜子叶颜色和球心色遗传关系的调查结果可以看出(表2):F1子叶全部表现为黄色,球心色也全部表现为普通心色;F2黄色子叶和橘红子叶的分离比是3∶1,普通心色和橘红心色的分离比也是3∶1;BC1F1子叶颜色的分离比与球心色的分离比一样,都是1∶1。而且子叶颜色与球心色一一对应,即橘红子叶的植株球心色也是橘红色,黄色子叶的植株球心色是普通心色。这说明橘红子叶颜色和橘红心色的遗传由同1 对基因控制,二者只是表现时间不同。
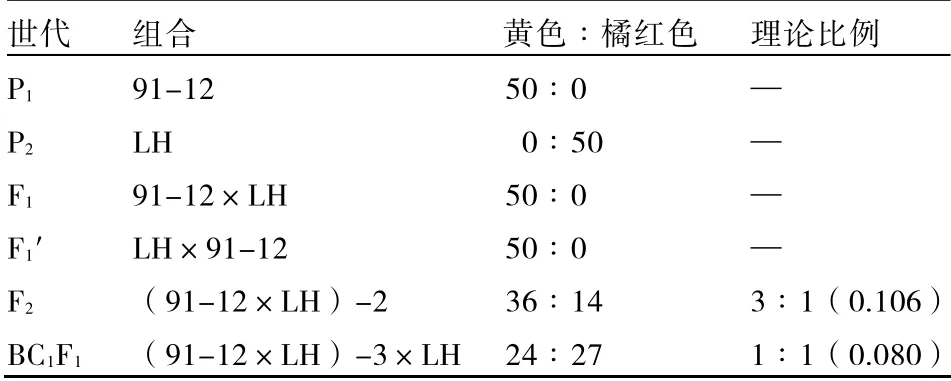
表1 P1、P2、F1、F1′、F2和BC1F1子叶颜色调查结果

表2 橘红心大白菜子叶颜色与球心色的遗传关系
子叶颜色与花色的遗传关系见表3,F1子叶全部为黄色,花色也全部表现为黄色,F2黄色子叶和橘红子叶的分离比与黄花和橘红花的分离比都是3∶1,BC1F1子叶颜色的分离比与花色的分离比也一样,都是1∶1,并且子叶颜色与花色的遗传一致,橘红子叶的植株花色是橘红色的。有研究指出橘红心大白菜花色与球心色是连锁遗传或一因多效(张德双 等,2003;冯辉 等,2009),球心色是橘红色,花色也是橘红色,反之也是如此。本试验获得相同的研究结果。由以上可知,橘红子叶、橘红心和橘红花三者表现为由同1 对隐性基因控制的遗传,属于一因多效。在大白菜试验周期中,橘红色出现的先后顺序为橘红子叶→橘红心→橘红花,橘红子叶出现的最早。

表3 橘红心大白菜子叶颜色与花色的遗传关系
2.2 橘红心大白菜子叶颜色在不同环境条件下的稳定性
室温条件下,光照处理与遮光处理大白菜幼苗子叶颜色差异明显(表4)。光照处理下,P1、P2、F1、F2和BC1F1幼苗子叶颜色相同,都表现为绿色。这是由于光是植物叶绿素合成的一个主要条件,光照条件下子叶合成的叶绿素的量大于类胡萝卜素的量,所以子叶都表现为绿色。遮光处理时子叶叶绿素合成受阻,P1、P2、F1、F2和BC1F1的子叶均表现为黄色。光照2 h 后,P1、F1子叶颜色依然是黄色,P2表现为橘红色,F2和BC1F1子叶颜色表现为黄色和橘红色的分离现象,分离比与橘红心子叶颜色的遗传规律相符合。
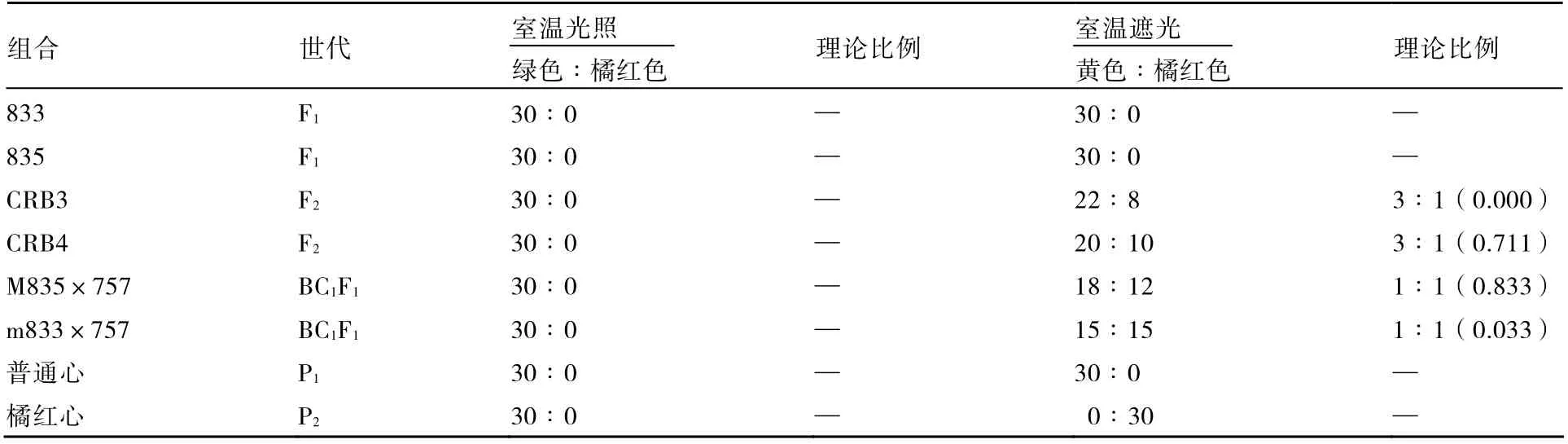
表4 光照2 h 后室温光照和室温遮光处理对大白菜子叶颜色的影响
低温条件下,遮光和光照处理结束时幼苗子叶颜色均表现为黄色,经过2 h 光照后,普通心P1和F1子叶仍然表现为黄色,橘红心P2子叶表现为橘红色,F2和BC1F1子叶颜色分别表现黄色∶橘红色为3∶1 和1∶1 的分离比例(表5)。低温遮光处理下橘红子叶颜色比低温光照处理下的橘红子叶稍深,低温光照处理的子叶橘红色与常温遮光处理的颜色相似。
另外,大白菜种子无论催芽24 h 还是催芽48 h 后进行低温遮光处理,处理结束后光照2 h,子叶颜色的分离表现一致(表 6),这说明催芽时间不同对橘红心大白菜子叶颜色的表现没有影响。
由以上可知,橘红心大白菜子叶颜色(橘红色)虽然在不同条件下表现出一定的深浅差异,但是和普通心大白菜子叶颜色(黄色)差异明显,可以很容易地区分出来。因此橘红心大白菜子叶颜色可以像其花色一样,作为一种理想形态标记来使用,特别是在转育复等位基因雄性不育系时可以减少大量的调查群体。在以橘红色子叶作为橘红心大白菜的形态标记时,以催芽48 h、低温、遮光条件下效果最为显著。
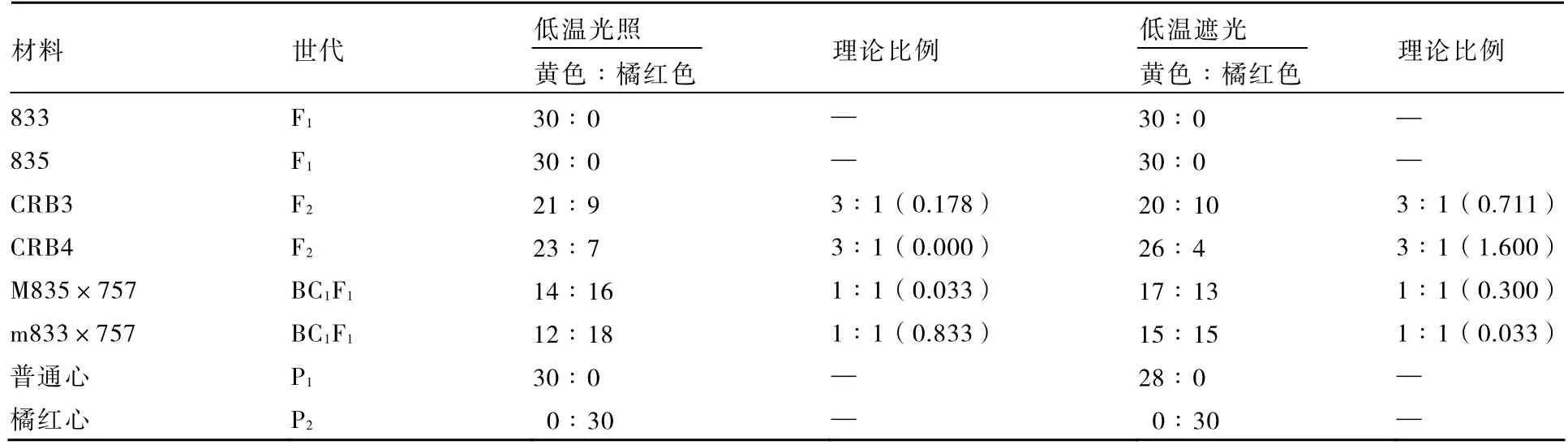
表5 光照2 h 后低温光照和低温遮光处理对大白菜子叶颜色的影响
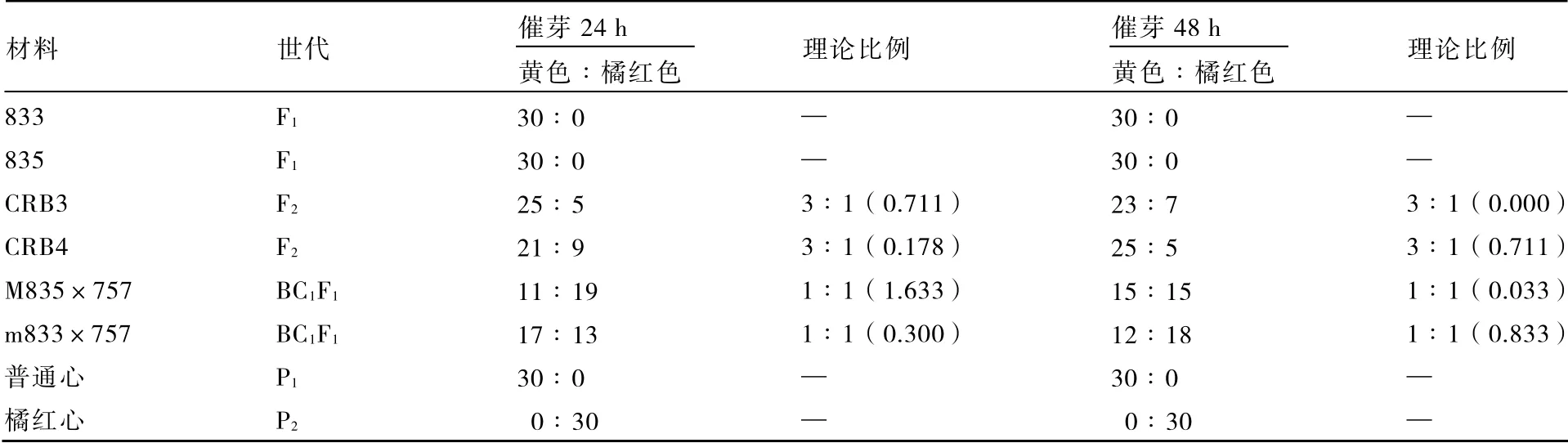
表6 低温遮光条件下不同催芽时间对大白菜子叶颜色的影响
3 结论与讨论
在苗期通过观察标记性状的存在与否保留理想植株,直观准确,简便快速,具有DNA 分子标记鉴定技术无法比拟的优越性,非常适合在我国相对落后的农业技术水平下推广(马双武,2011)。分子标记技术准确性高,不受环境的影响,无器官发育时期的特异性,利用橘红心大白菜SCAR 标记可以鉴定种子纯度(王国臣 等,2007)。然而,分子标记辅助选择需要建立饱和的分子标记图谱,建立分子标记与目标基因的连锁关系和检测方法的自动化(王琦,2006)。目前大白菜球色育种上分子标记的应用尚处于探索阶段(张立志和张鲁刚,2011)。
在本试验中,遮光处理结束后所有材料子叶颜色均为黄色,光照2 h 后,橘红心大白菜和普通心大白菜子叶颜色表现出明显差异。这可能是因为光照时间较短,普通心大白菜子叶叶绿素还没有来得及合成或合成量很少(孟繁静 等,1995),所以表现为黄色。但是光对类胡萝卜素合成的调节机制非常复杂,例如番茄幼苗脱黄化的过程中,体内的类胡萝卜素含量可增长几倍(惠伯棣,2005),橘红心大白菜子叶颜色在光照2 h 后由黄色转变为橘红色可能是由于体内叶绿素还没有合成、而类胡萝卜素含量迅速增加的缘故。而室温光照处理结束时橘红心大白菜子叶颜色为绿色、低温光照处理结束时橘红心大白菜子叶颜色为黄色可能是低温光照处理的温度低、光强弱,叶绿素难以合成的缘故。
本试验通过对橘红心大白菜自交系和普通心大白菜自交系的杂交、自交和回交后代子叶颜色遗传规律和稳定性的研究得出如下结论。
①橘红心大白菜子叶颜色橘红色对普通心大白菜子叶颜色黄色表现为1 对基因控制的遗传,橘红色对黄色表现为隐性,并且子叶颜色与球心色和花色相对应,三者的遗传属于一因多效。
②自然光照处理下,子叶叶绿素合成量多于类胡萝卜素合成量,橘红心大白菜子叶颜色与普通心大白菜子叶颜色相同,均为绿色。在弱光(200 lx)处理后光照2 h,叶绿素还没有合成或含量很低,普通心大白菜子叶表现为黄色,橘红心大白菜子叶表现为橘红色。
③无论常温还是低温,遮光处理结束后光照2 h,橘红心大白菜子叶表现为橘红色,普通心大白菜子叶表现为黄色,橘红色与黄色差异明显,容易区分。
④在常温遮光条件下橘红心大白菜子叶颜色表现稳定,低温遮光条件下橘红心大白菜子叶的橘红色更加明显,催芽时间不同不影响橘红心大白菜子叶颜色的表现。不仅如此,橘红子叶在子叶期就可以观察到,即在幼苗初期就可以鉴定,转育复等位基因雄性不育系时可以大量减少后期的调查群体。因此可以说,橘红心大白菜子叶颜色是球心色的一个理想的形态标记性状,利用这一性状在育种过程中可以进行早期筛选,加快育种进程。
冯辉,姜楠,李成彧.2009.橘红心大白菜核基因雄性不育系转育方法研究.中国蔬菜,(12):48-52.
惠伯棣.2005.类胡萝卜素化学及生物化学.北京:中国轻工业出版社:53-54.
马双武.2011.苗期标记性状在我国作物育种及种子纯度鉴定上的研究应用进展.植物遗传资源学报,12(2):297-300.
孟繁静,刘道宏,苏业瑜.1995.植物生理生化.北京:中国农业出版社:152,156-158.
涂金星,傅廷栋.2001.甘蓝型油菜几个苗期形态标记的遗传.华中农业大学学报,20(4):318-320.
王国臣,张凤兰,余阳俊,张德双,赵岫云.2007.与大白菜橘红心基因紧密连锁的SCAR 标记.园艺学报,34(1):217-220.
王琦.2006.大白菜橙色叶球基因分子标记及杂种纯度鉴定研究〔硕士论文〕.杨凌:西北农林科技大学.
吴新杰,陈凤祥,胡宝成,李强生,侯树敏,费维新,黄晓荣,江莹芬.2005.甘蓝型油菜形态标记性状研究进展.中国农学通报,21(12):128-131.
余阳俊,陈广,段建雄.1993.大白菜球色的遗传.北京农业科学,11(5):18-19.
张德双,张凤兰,徐家炳.2003.大白菜花色和球色遗传规律的研究.华北农学报,18(2):81-84.
张立志,张鲁刚.2011.大白菜球叶颜色育种研究进展.中国瓜菜,24(2):38-41.
钟淮钦,李富生,杨清辉.2005.甘蔗杂种真实性鉴定研究综述.中国农学通报,21(6):390-394.
猜你喜欢
西北植物学报(2022年11期)2023-01-04
Chinese Physics B(2022年11期)2022-11-21
中学生数理化(高中版.高考数学)(2022年2期)2022-04-26
智慧少年·故事叮当(2019年2期)2019-06-11
中学生数理化(高中版.高二数学)(2018年1期)2018-02-26
创新作文(5-6年级)(2017年6期)2017-12-25
作文周刊·小学一年级版(2017年38期)2017-11-11
植物资源与环境学报(2017年3期)2017-10-12
中学数学杂志(高中版)(2017年4期)2017-07-27
中国公路(2017年9期)2017-07-25
