
广西新分离流行性乙型脑炎病毒GP0722株全基因组分子特性研究*
2012-01-24 07:44闭福银谢艺红杨进业
中国人兽共患病学报 2012年3期
闭福银,谢艺红,谭 毅,杨进业
广西新分离流行性乙型脑炎病毒GP0722株全基因组分子特性研究*
闭福银,谢艺红,谭 毅,杨进业
目的对广西新分离乙脑病毒GP0722株进行全基因序列测定和分析,了解其基因组结构及毒力特征。方法应用乙脑病毒全基因组扩增引物进行RT-PCR扩增,PCR产物直接测序,拼接后得到全基因序列。应用Clustal X(1.8)、DNASTAR、Mega 4.1等生物软件进行核苷酸序列及氨基酸序列分析和病毒的系统进化分析。结果广西新分离乙脑病毒GP0722全基因长10 965个核苷酸,从97到10 395位编码一个开放阅读框,编码3 432个氨基酸,与目前使用的减毒活疫苗株SA-14-14-2株比较,只有88.9%的核苷酸同源性,97.6%的氨基酸同源性,全基因组共存在1 222个核苷酸差异,83个氨基酸差异。与GenBank中选择的21株乙脑病毒全基因序列比较发现,其核苷酸总体差异率为0.9%~18.8%,氨基酸总体差异率为0.1%~5.2%。通过Pr M/C区段、E区段、3′NTR区段和全基因序列进行系统进化分析显示该毒株属于基因1型乙脑病毒。结论新分离的乙脑病毒GP0722株属于基因1型,与JEV/sw/Mie/40/2004进化关系最近,与疫苗株SA-14-14-2相比关键位点氨基酸未见变异,现行使用的疫苗仍能保护GP0722引起的感染。
乙型脑炎病毒;基因;种系发生
流行性乙型脑炎(Japanese encephalitis,JE,乙脑),是由乙脑病毒(Japanese encephalitis virus,JEV)感染引起的经蚊虫传播的人兽共患急性传染病。乙脑患者的病死率高,幸存者中约30% 的人会留有不同程度的中枢神经系统后遗症[1-2]。
乙脑属于黄病毒科黄病毒属,基因组为约11kb的单股正链RNA,5′端有1型帽子结构,3′端不含poly尾。基因组只有一个开放读码框,主要编码3个结构蛋白(衣壳蛋白、膜蛋白、囊膜蛋白E)和7个非结构蛋白(NS1、NS2a、NS2b、NS3、NS4a、NS4b、NS5),基因组5′末端包括95个核苷酸,3′末端包括586个核苷酸[3]。
乙脑主要在中国、日本和越南等亚洲国家和地区流行,近年来乙脑的流行区域不断扩大[4],目前全世界约20余个国家存在乙脑流行,我国为乙脑的高发区,平均年发病数约占全世界乙脑病例的80%以上[5]。
广西从1950年开始有乙脑病例报告,报告显示广西20世纪60年代初期至70年代中期乙脑发病率呈上升并维持在较高水平,至90年代后开始呈下降趋势[6-7],特别是近年来广西将乙脑纳入计划免疫管理,疫情进一步得到控制,但每年还有几十例的病例报告,因此,了解广西乙脑的分子生物学特征,对乙脑防控非常必要。
1 材料和方法
1.1 毒株背景2007年从桂平三带喙库蚊中分离到乙脑病毒GP0722株。
1.2 病毒RNA的提取及全基因组序列测定 取病毒培养物,High Pure Viral RNA Kit(Roche,德国)试剂盒提取核酸。引物序列参照文献[8],由上海生工生物科技有限公司合成,RT-PCR试剂使用QIAGEN OneStep RT-PCR Kit(Qiagen,德国)。
1.3 病毒核苷酸及氨基酸序列分析 使用DNAstar软件包中的Seq Man对序列片段进行拼接、编辑、校正。碱基配对由Clustal X1.8软件完成,核苷酸及氨基酸序列同源性和差异度分析由DNAstar软件包中的Meg Align软件完成。
1.4 病毒株的进化分析 对GP0722株及目前GenBank登录的、覆盖基因1~4型的JEV全基因组序列,由Clustal X1.8软件进行病毒核苷酸序列联配,选择莫雷山谷热病毒 MVE为外群,用MEGA4.1软件中的Neighbor-Joining方法进行序列分析,获得最终的系统进化树。
2 结 果
2.1 GP0722株全基因组序列 将所得的PCR产物进行双向测序,拼接后得到病毒的全基因序列。病毒基因组全长10 965个核苷酸,5′非编码区包括96个核苷酸,开放阅读框从97至10 395,共10 296个核苷酸,编码3 432个氨基酸,3′非编码区包括570个核苷酸。
2.2 GP0722株与减毒活疫苗株序列比较分析与减毒活疫苗株SA14-14-2相比,全基因组核苷酸差异率为11.14%,共存在1 222个核苷酸差异,氨基酸差异率为2.42%,存在83个氨基酸差异。其中非结构基因NS2的核苷酸的差异率最高为12.53%,5′端非编码区的差异率最低为3.13%;非结构基因NS2的氨基酸差异率最高为3.69%,非结构基因NS3的氨基酸差异率最低为1.45%(表1)。在结构基因中共有24个氨基酸的差异,其中7个在衣壳基因,3个在膜基因,14在囊膜基因(表2)。
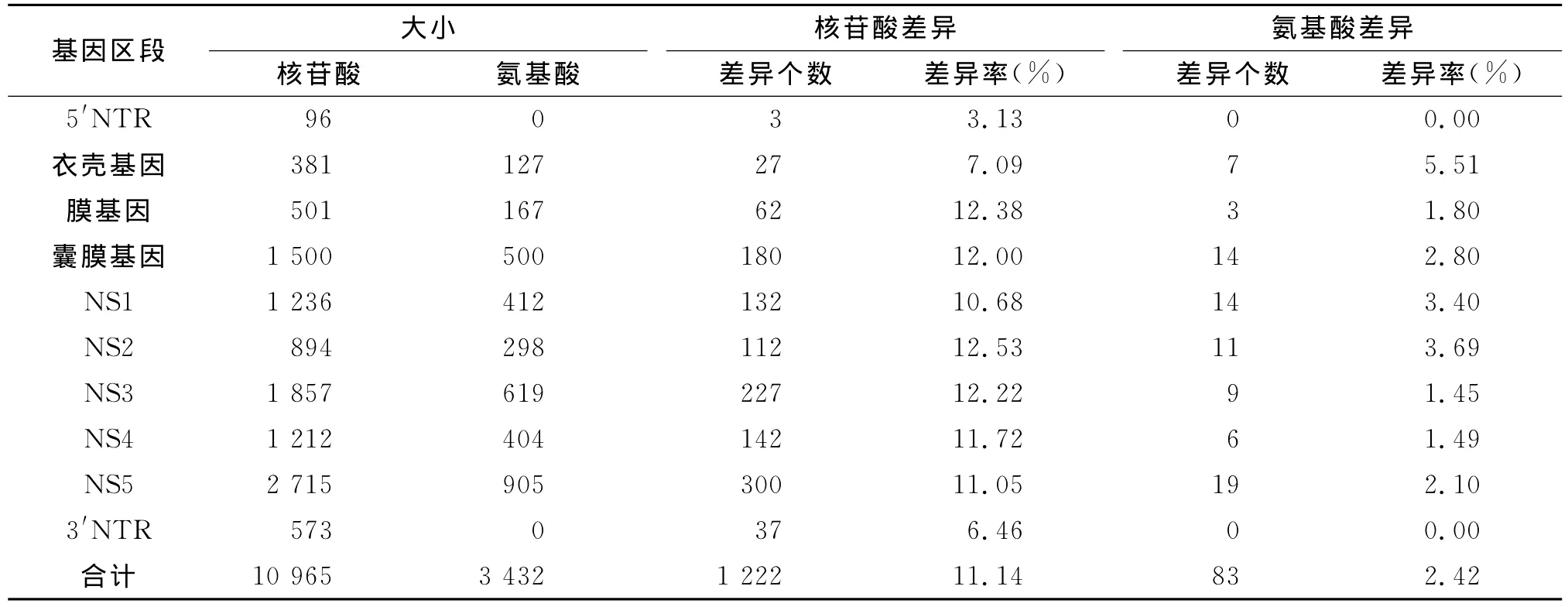
表1 乙脑毒株GP0722与减毒活疫苗株SA14-14-2之间基因组比较分析Tab.1 Genome sequence analysis of the GP0722 compared with vaccine strain SA14-14-2
2.3 GP0722株与其他乙脑毒株全基因组序列核苷酸及氨基酸差异分析 将GP0722株与GenBank中选取的其他国家和地区分离的覆盖4个基因型别的21株乙脑毒株的全基因核苷酸及氨基酸序列进行同源性分析,GP0722株与其他毒株的核苷酸总体差异率为0.9%~18.8%,氨基酸总体差异率为0.1%~5.2%。其中 GP0722株与JEV/sw/Mie/40/2004株的核苷酸差异率最低为0.9%,与JKT6468株核苷酸差异率最高为18.8%;与JEV/sw/Mie/40/2004氨基酸同源性最高为99.9%,与JKT6468氨基酸同源性最低为94.8%。
2.4 系统进化分析 将GP0722株与21株乙脑病毒株的Pr M/C区段、E基因区段、3′NTR区段及全基因序列进行核苷酸联配后,进行进化分析,选择1株莫雷山谷热病毒作为外群。4种分析方法均可以看到明显的4个基因型别,其中基因1型7株,基因2型1株,基因3型13株,基因4型1株,4种分析方法均显示新分离的乙脑病毒GP0722株在基因1型中(图1)。
3 讨 论
自从乙型脑炎被证实在中国流行以来,国内各地分离到了大量的乙脑病毒株并进行了一定的研究,但大多集中在于高变异的Pr M/C区段[9],基于不同的地理来源将乙脑病毒分为4个基因型。本研究对2007年在广西桂平市捕捉的三带喙库蚊标本中新分离的乙脑病毒株GP0722株进行全基因组序列测定,对病毒基因进行了Pr M/C区段、E区段、3′NTR区段和全基因序列进化分析,结果完全一致。
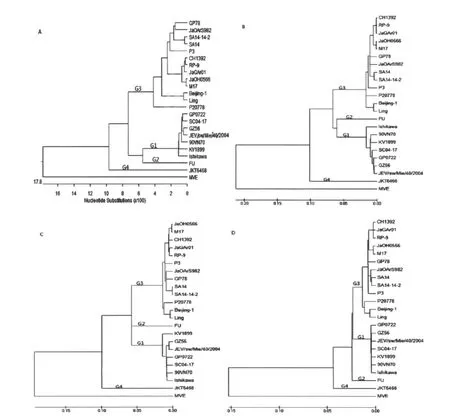
图1 分离自不同国家的22株乙脑病毒的全基因序列系统进化分析MAV:莫雷山谷热病毒缩写,作为进化分析的外群;A:全基因;B:E区段;C:Pr M/C;D:3′NTR区段Fig.1 Phylogenetic tree based on the full-length genome of genetic relationships among 22 JE virus strains MVE sered as outgroup;A:Full-length;B:E;C:Pr M/C;D:3′NTR

表2 乙脑病毒GP0722株与减毒活疫苗株SA14-14-2结构蛋白氨基酸差异分析Tab.2 Amino acid substitutions in JEV stains(GP0722)structural proteins compared with vaccine strain SA14-14-2
减毒活疫苗株SA14-14-2是目前国内扩大免疫规划广泛使用的疫苗株,分析比较新分离株与其全基因组序列对了解疫苗对该地区乙脑流行的保护具有重要的意义,E蛋白是流行性乙型脑炎的主要抗原成分,具有特异性中和及血凝抑制抗原决定簇,E蛋白结构域Ⅲ 是重要的抗原表位,其中主要集中在E337-345,E377-382 和 E397-403 这 3 个 区 域[10]。本次研究结果显示GP0722株和疫苗株SA14-14-2在这3个区域完全一致,本研究结果从分子角度证实了减毒活疫苗株SA14-14-2株能保护GP0722株引起的感染。另一方面,对TBE病毒的研究发现E306是决定黄病毒对鼠脑神经侵入力的关键位点[11];对病毒神经侵入力研究认为,E52和E270位氨基酸可影响病毒进入宿主细胞,从而降低病毒神经侵入力[12-13];对病毒神经毒力的研究还认为,当乙脑病毒E138位的氨基酸由谷氨酸替换成赖氨酸后病毒的神经毒力会有明显的降低[14];通过比较发现,GP0722的 E52、E270和 E306与 SA14-14-2一致,E138位点氨基酸为谷氨酸,说明GP0722神经毒力未发生明显降低。
广西位于我国西南端,全年平均气温16℃~22.7℃之间,气候湿润,雨水丰富,适合各种媒介昆虫的繁殖,并且广西与虫媒病毒高发的地区广东、云南、越南等省毗邻,随着南宁成为东盟博览会永久举办地,各地区的经贸合作频繁,以及广西旅游业的快速发展,广西存在着各种虫媒病毒病暴发的风险相当高。因此,加强广西虫媒病毒的研究非常有必要。本研究对2007年在桂平市采集的蚊虫标本中分离的乙脑病毒进行的全基因组序列测定,分析了其分子生物学特性,为预防和控制乙脑在当地的流行提供了分子流行病的科学依据。
[1]Gould EA,Solomon T.Pathogenic flaviviruses[J].Lancent,2008,371:500-509.
[2]Solomon T,Ni H,David W C,et al.Origin and evolution of Japanese encephalitis virus in southeast Asia[J].J Virol,2003,77:3091-3098.
[3]自登云,陈伯权,俞永新.虫媒病毒与虫媒病毒病[M].昆明:云南科技出版社,1995:157.
[4]梁国栋.加强中国病毒性脑炎的检测与研究[J].中华实验和临床病毒学杂志,2006,20(3):201-202.
[5]王环宇,梁国栋.中国分离乙脑病毒与灭活疫苗株(P3株)E基因差异分析[J].中华实验和临床病毒学杂志,2006,20(1):56-60.
[6]谭毅,谢艺红,周开姣,等.1990-2007年广西流行性乙型脑炎流行病学特征分析[J].应用预防医学,2008,14(6):323-325.
[7]孟军,杨进业,吴秀玲,等.广西1992-2006年流行性乙型脑炎流行特征分析[J].中国热带医学,2007,7(8):1422-1423.
[8]潘晓玲,梁国栋.乙脑病毒基因Ⅰ、Ⅲ型特异性全基因组引物[J].中华实验和临床病毒学杂志,2009,23(4):254-256.
[9]Chen WR,Tesh RB,Rico-Hesse R.Genetic variation of Japanese encephalitis virus in nature[J].J Gen Virol,1990,71:2915-2922.
[10]Lobigs M,Uaha R.Host cell selection of murray valley encephalitis virus variants altered at an RGD sequence in the envelope protein and in mouse virulence[J].Virology,1990,176:587-595.
[11]Rey FA,Heinz FX,Mandl C,et al.The envelope glycoprotein from tickborne encephalitis virus at 2 A resolution[J].Nature,1995,375:291-298.
[12]Hasegawa H,Yoshida M,Shiosaka T,et al.Mutation in the envelope protein of Japanese encephalitis virus affect entry into cultured cells and virulence in mice[J].Virology,1992,191:158-165.
[13]Cecilia D,Gould EA.Nucleotide changes responsible for loss of neuroinvasiveness in Japanese encephalitis virus neutralizationresistant mutants[J].Virology,1991,181:70-77.
[14]Ni H,Chang GJ,Xie H,et al.Molecular basis of attenuation of neurovirulence of wild-type Japanese encephalitis virus strain SA14[J].J Gen Virol,1995,76:409-413.
Molecular characteristics of full-length genome of Japanese encephalitis virus strain(GP0722)newly isolated in Guangxi province
BI Fu-yin,XIE Yi-hong,TAN Yi,YANG Jin-ye
(Guangxi Zhuang Autonomous Region Center for Disease Control and Prevention,Nanning 530028,China)
In order to sequence and analyze the complete nucleotide sequence of Japanese encephalitis virus(JEV)strain GP0722 newly isolated in Guangxi province,and to provide information for the genomic structure and characteristics of virulence.RT-PCR was used to amplify the sequenced fragments directly and bioinformatics softwares were used to analyze the nucleotide,amino acid and phylogenetic trees of the strain including ClustalX(1.8),DNAStar,Mega(4.1).The result of sequence analysis showed that the genome of GP0722 strain was 10 965 nucleotide long.An open reading frame from 97 to 10 395 was found capable of coding for a 3 432 amino acid polyprotein.Compared with the live attenuated vaccine strain SA14-14-2 in China,there were 1 222 nucleotide differences and 83 amino acid divergence.Comparison of the complete genome sequences of different JEV showed a 0.9%-18.8%nucleotide sequence divergence among them,which resulted in 0.1%-5.2%amino acid sequence divergence.Phylogenetic analysis through Pr M/C,E,3′NTR and full-length genome showed that the GP0722 strain belonged to genotype 1.In conclusion,the GP0722 strain isolated in Guangxi belonged to genotype 1 and is close to the JEV/sw/Mie/40/2004.There are no mutations.Vaccination can protect human from the infection of GP0722.
Japanese encephalitis virus;genes;phylogeny
R373
A
1002-2694(2012)03-0270-04
*广西壮族自治区卫生厅计划课题(No.Z2008204)
闭福银,Email:bfy1980@sina.com
广西壮族自治区疾病预防控制中心,南宁 530028
2011-10-11
2012-01-16
猜你喜欢
世界科学技术-中医药现代化(2022年3期)2022-08-22
肝博士(2022年3期)2022-06-30
云南画报(2021年11期)2022-01-18
铁道通信信号(2021年6期)2021-07-08
科技视界(2020年8期)2020-05-18
健康博览(2019年8期)2019-09-12
妈妈宝宝(2019年6期)2019-07-02
铁道通信信号(2019年3期)2019-04-25
中国饲料(2019年19期)2019-03-25
