
不同立木级木荷对低温胁迫的生理响应研究
2012-01-19 03:28柳翼飞王小德曹俊卓张晶月
浙江农业科学 2012年5期
柳翼飞,王小德,黄 可,曹俊卓,张晶月
(浙江农林大学 风景园林与建筑学院,浙江 临安 311300)
木荷 (Schima superba),为亚热带树种,具有许多优良性状,集用材、抗灾和观赏于一体,是近年来大力发展的珍贵乡土树种,在植被恢复及林业生产上得到越来越广泛的应用。低温伤害使细胞膜受损,细胞膜相对透性变大[1],导致细胞内的电解质外泄。因此,电解质外渗率的变化能反映组织受伤害程度和植物的抗寒性大小,实验中通常用逆境伤害前后质膜的相对电导率来直观的表示膜内电解质外渗率[2]。低温下,植物体可通过保护性酶如超氧化物歧化酶 (SOD)、过氧化物酶 (POD)等,清除活性氧自由基[3]、积累大量渗透调节物质 (可溶性蛋白、可溶性糖等)[4]调节细胞渗透压等方式减轻低温对植物的伤害。很多实验证实[5-8],SOD-POD的活性水平可以间接反映植物抗寒能力的高低,而可溶性蛋白、可溶性糖含量的增加有利于植物抗寒性的提高。
本研究以5种立木级木荷为试验材料,通过抗寒性生理指标的测定对木荷的抗寒机制进行初步探讨,为木荷的南树北扩以及栽培养护提供理论依据。
1 材料与方法
1.1 材料
试验的木荷选自浙江农林大学植物园。参照立木级的划分标准,每个立木级选择平均标准左右、具有代表性的木荷植株,生长状况基本相同。Ⅰ级:划分标准h<0.33 m,试材规格0.2 m <h<0.3 m。Ⅱ级:划分标准 d<2.5 cm;h>0.33 m,试材规格1 cm <d<2 cm;1.5 m<h<2.5 m。Ⅲ级:划分标准d=2.5~7.5 cm,试材规格d=4~7 cm。Ⅳ级:划分标准d=7.5~22.5 cm,试材规格d =25~ <30 cm。Ⅴ级:划分标准 d>22.5 cm,试材规格d=4~7 cm。进行人工模拟低温胁迫处理的叶片系从树冠外围中部采取的新鲜叶片,要求生长良好,长势基本一致,无病虫害。
1.2 方法
新鲜叶片分别用自来水、蒸馏水冲洗,用吸水纸吸干水分。每立木级叶片各分出6组置于密封塑料袋内,将其中1组置于25℃室温做对照,其他5组放入冷却循环水浴仪。样品于15℃预冷5 h,降温至10℃预冷7 h。再从10℃开始进行降温,每次降温5℃,即10℃→5℃,在5℃停留2 h后,取出第1组样品,然后继续降温至0℃,停留2 h后再取出第2组样品,依此类推,直至温度降至-15℃。将低温处理结束后取出的样品放置于4℃冰箱内解冻恢复2 h后,待测。
1.3 测定指标及方法
用电导法测相对电导率;氮蓝四唑法测SOD活性;蒽酮比色法测可溶性糖含量;考马斯亮蓝法测可溶性蛋白含量;愈创木酚法测POD活性[9-10]。
2 结果与分析
2.1 电导率
由图1可以看出,5种立木级木荷的相对电导率都随处理温度的下降呈明显的“S”型曲线,且均在-5℃时值最小。其中,Ⅱ级、Ⅲ级在5℃降至0℃时,相对电导率小幅升高,随着温度降至-5℃,略有下降,在 -5℃降至最低,后持续升高;Ⅰ级、Ⅳ级、Ⅴ级变化趋势一致,轻度低温胁迫 (5~0℃)时,相对电导率缓慢降低,在-5℃时降至最低,后快速升高。
对低温处理下不同立木级木荷叶片的相对电导率进行多重比较 (表1),表明,Ⅱ级、Ⅳ级的相对电导率没有显著差异,而其他立木级间相对电导率差异显著;Ⅲ级木荷的相对电导率与其他4种立木级木荷的相对电导率存在极显著差异,Ⅰ级与Ⅴ级没有极显著差异。
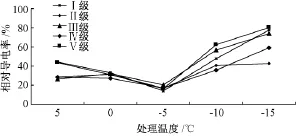
图1 低温处理对5种立木级木荷叶片相对电导率的影响
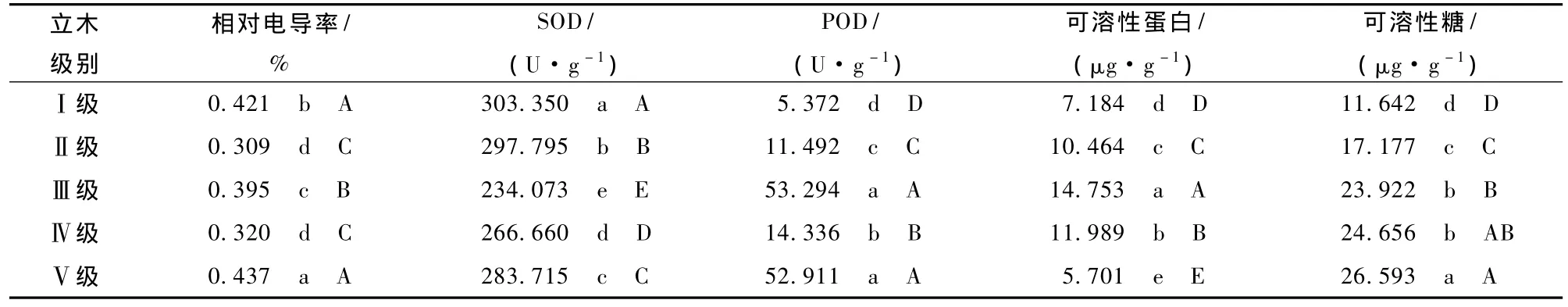
表1 低温处理后木荷不同立木级间抗寒指标的多重比较
对不同低温处理下木荷叶片的相对电导率进行多重比较 (表2)表明,所有处理温度下木荷叶片的相对电导率与常温对照相比都有极显著差异,不同的处理温度之间也存在极显著差异。
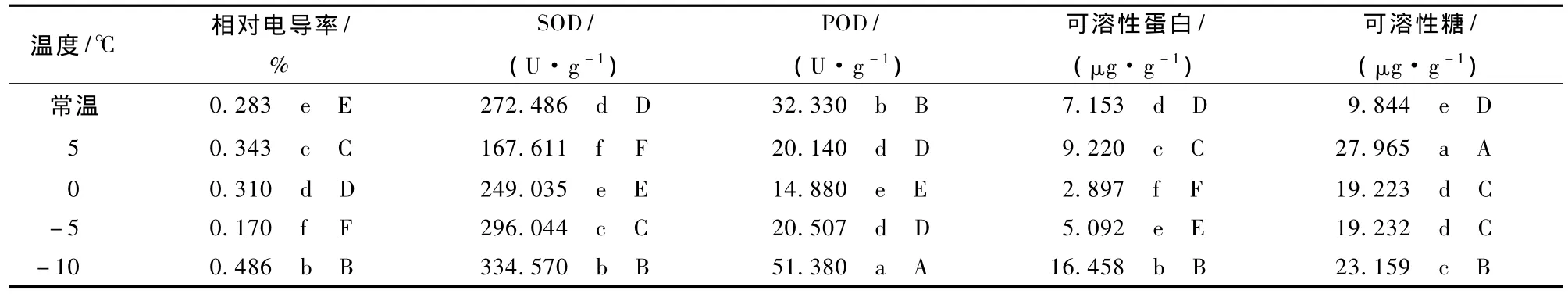
表2 不同温度处理木荷叶片抗寒性指标的多重比较
综合来看,木荷的细胞电解质外渗率随着温度的降低而升高,有时出现电解质外渗率降低的现象,说明木荷的抗寒机制可在一定程度上修复细胞膜系统,但是恢复作用有限,如果胁迫加重,并不能阻止电解质的外渗加剧。值得注意的是,在处理温度降至-10℃后,Ⅰ级木荷相对电导率增长趋于停滞,其他4种立木级木荷的相对电导率增长趋缓,可能是因为木荷的抗寒防卫作用渐趋极限,细胞膜发生不可逆的损伤,细胞膜内外渗透压趋于平衡所致。由此推测,如果温度继续降低,5种立木级木荷的细胞外渗率可能都会达到极值而不再增加。
2.2 超氧化物歧化酶 (SOD)活性
如图2所示,Ⅰ级木荷SOD活性随处理温度的降低,先升后降,于-10℃时达到峰值;Ⅱ级木荷SOD活性先升高,在-5℃略降后继续升高;Ⅲ级、Ⅳ级、Ⅴ级木荷的SOD活性均随处理温度的降低持续升高。低温处理后,5种立木级木荷的SOD活性差异极显著,Ⅰ级木荷的SOD活性极显著高于其他立木级,Ⅲ级木荷的SOD活性极显著低于其他立木级 (表1)。所有处理温度下木荷的SOD活性均与常温对照存在极显著差异,不同的处理温度之间也存在极显著差异 (表2)。
综合来看,木荷的SOD活性随处理温度的降低大体呈上升趋势,但是,升势缓慢,Ⅰ级木荷在低于-10℃时SOD活性甚至下降,说明木荷通过提高SOD活性来对抗低温胁迫是有限度的,低温胁迫严重到一定程度,SOD活性很可能开始下降。
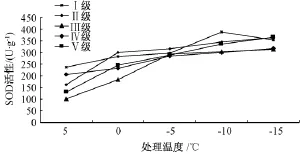
图2 低温处理对5种立木级木荷SOD活性的影响
2.3 POD活性
如图3所示,Ⅰ级、Ⅳ级、Ⅴ级木荷叶片的POD活性均随处理温度的降低先升后降,其中,Ⅰ级在-5℃达到峰值,Ⅳ级、Ⅴ级在-10℃达到峰值;Ⅱ级、Ⅲ级呈现“S”型变化,均在0℃时活性最低,-10℃活性最高。Ⅲ级、Ⅳ级木荷叶片的POD活性无显著差异,极显著高于其他立木级木荷;其他立木级间差异极显著,其中,Ⅰ级木荷叶片的POD活性极显著低于其他立木级 (表1)。低温处理的所有温度均与常温对照差异极显著,不同的处理温度之间也存在极显著差异(表2)。
综合来看,虽然拐点温度不尽相同,但是当温度降到一定程度时,5种立木级木荷的POD活性均出现由升转降的现象,说明轻度低温会诱导木荷提高POD活性,但是,严重低温会导致木荷的POD活性降低。
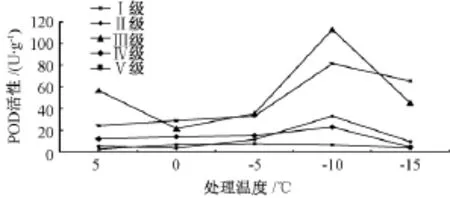
图3 低温处理对5种立木级木荷POD活性的影响
2.4 可溶性蛋白含量
如图4所示,5种立木级木荷可溶性蛋白含量对低温胁迫的响应呈较一致的变化趋势,大体上先降后升,均在0℃时取得最小值。不同立木级木荷可溶性蛋白含量差异极显著;Ⅲ级木荷的可溶性蛋白含量极显著高于其他立木级,Ⅴ级木荷叶片的可溶性蛋白含量极显著低于其他立木级 (表1)。低温处理的各个温度均与常温对照的可溶性蛋白含量差异极显著,且不同处理温度之间的可溶性蛋白含量也差异极显著 (表2)。
综合来看,随处理温度的降低,木荷叶片的可溶性蛋白含量均先减少再增加。前期的低温处理中,可溶性蛋白的减少,说明木荷通过增加蛋白质含量对抗低温的机制需要一定低温的诱导,在温度未达到需要的低温时,可溶性蛋白含量是减少的。
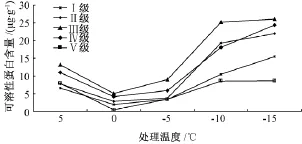
图4 低温处理对5种立木级木荷可溶性蛋白含量的影响
2.5 可溶性糖含量
如图5所示,Ⅰ级、Ⅱ级木荷可溶性糖含量随温度的降低缓慢减少,在-10℃时达到最小值后略有回升;Ⅲ级、Ⅳ级、Ⅴ级的可溶性糖含量以0℃为拐点,先降后升。经低温处理,Ⅲ级与Ⅳ级可溶性糖含量差异不显著;而其他立木级间差异显著,其中,Ⅳ级与Ⅴ级、Ⅳ级与Ⅲ级无极显著差异,而与Ⅰ级、Ⅱ级差异极显著;Ⅰ级的可溶性糖含量极显著低于其他立木级 (表1)。所有处理温度的可溶性糖含量均极显著大于常温对照。0℃与-5℃时,可溶性糖含量无显著差异 (表2)。
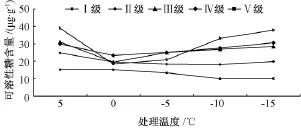
图5 低温处理对5种立木级木荷可溶性糖含量的影响
综合来看,在轻度低温胁迫 (常温 -5℃)时,5种立木级木荷的可溶性糖含量都大幅增高,随着温度继续降低,变化趋缓,增减均不显著。说明,木荷在轻度低温胁迫时,积累可溶性糖的能力较强,而随着温度进一步降低,积累可溶性糖的机制受阻。
3 小结
与常温对照相比,低温处理下5种立木级木荷的SOD活性、POD活性、可溶性蛋白含量整体呈上升趋势,可溶性糖含量均显著高于常温对照,但随着温度的变化不显著。Ⅰ级木荷叶片相对电导率最大,SOD活性最高,POD活性、可溶性糖含量最低;Ⅱ级木荷叶片相对电导率最小;Ⅲ级POD活性、可溶性蛋白含量最高,SOD活性最低;Ⅴ级可溶性糖含量最高,可溶性蛋白含量最低。说明不同立木级木荷对低温胁迫的生理响应差异极大,各有侧重。
综合来看,随处理温度的降低,木荷SOD活性与POD活性变化幅度差异很大,但变化趋势较一致,说明在木荷的抗寒机制中,SOD与POD是协同作用的。轻度低温胁迫时,木荷通过积累可溶性糖来提高抗寒性,低温胁迫较严重时,则通过可溶性蛋白的积累来提高抗寒性。
木荷不同的生理指标对低温胁迫的响应差异显著,不同立木级同一生理指标对低温胁迫的响应呈现较一致的变化规律,但变化幅度差别很大。说明不同立木级木荷的抗寒防卫机制相同,但单因素的调节能力因立木级的不同而存在显著差异。所有的指标均在低温胁迫到一定程度时,变化趋缓。说明木荷的抗寒机能有其极限,只能在一定程度上减轻低温对植物体的伤害。
[1] 彭筱娜,易自力,蒋建雄.植物抗寒性研究进展 [J].生物技术通报,2007(4):123-126.
[2] 牛立新,贺普超.用电导法不同计量单位鉴定葡萄抗寒性研究 [J].果树科学,1989(3):159-164.
[4] 江福英,李延,翁伯琦.植物低温胁迫及其抗性生理[J].福建农业学报,2002,17(3):190-195.
[3] 刘从霞.4种李属彩叶植物抗寒生理研究 [D].保定:河北农业大学,2007.
[5] 薛建辉,苏敬,刘金根,等.5个常绿阔叶园林树种对低温变化的生理响应 [J].南京林业大学学报:自然科学版,2009,33(4):38-42.
[6] 相昆,张美勇,徐颖,等.不同核桃品种耐寒特性综合评价 [J].应用生态学报,2011,22(9):2325-2330.
[7] 贺磊,游凯,李远芳,等.5种典型热带引种园林植物幼苗抗寒性研究 [J].中南林业科技大学学报,2011,31(2):65-71.
[8] 冯献宾,董倩,李旭新,等.黄连木和黄山栾树的抗寒性[J].应用生态学报,2011,22(5):1141-1146.
[9] 高俊凤.植物生理学实验指导 [M].北京:高等教育出版社,2006.
[10] 邹奇.植物生理学实验指导 [M].北京:中国农业出版社,2000.
猜你喜欢
石油库与加油站(2022年3期)2022-08-30
林业科学(2022年5期)2022-08-20
浙江农林大学学报(2022年4期)2022-08-18
中国林副特产(2022年4期)2022-08-08
卫星应用(2022年3期)2022-05-23
散文诗世界(2022年5期)2022-05-23
新农民(2021年21期)2021-09-17
矿冶工程(2021年4期)2021-09-15
花卉(2021年2期)2021-01-21
世界有色金属(2020年15期)2020-10-10
