
青藏高原东缘不同草地类型土壤酶活性研究
2012-01-02 02:48索南吉谈嫣蓉朱炜歆顾振宽杜国祯
草业学报 2012年4期
索南吉,谈嫣蓉,朱炜歆,顾振宽,杜国祯
(兰州大学干旱与草地生态教育部重点实验室,甘肃 兰州730000)
土壤酶主要来自于土壤微生物代谢过程,也能由土壤动物、植物残体分解产生[1]。土壤酶作为土壤系统的组分之一,是生态系统的生物催化剂,在土壤物质循环和能量转化过程中起着重要作用。它们参与包括土壤生物化学过程在内的物质循环,既是土壤有机物转化的执行者,又是植物营养元素的活性库[2,3]。20世纪80年代中期以后,土壤酶学的研究已经超越了经典土壤学的研究范畴,在几乎所有的陆地生态系统中,土壤酶活性的检测似乎成了必不可少的测定指标[4,5]。对退化土壤生态系统的研究发现,土壤酶活性的高低不仅与土壤生态系统的退化有关,而且与土壤类型、植被特征、微生物数量、土壤动物类群及数量有关[6]。由于土壤酶活性与土壤生物、土壤理化性质和环境条件密切相关[7],因而土壤酶活性对环境扰动的响应、根际土壤酶功能的重要性、土壤酶研究技术以及土壤酶作为土壤质量的指标等成为主攻方向[8-10]。
青藏高原因海拔高和气候寒冷,被认为是气候变化的敏感区和脆弱区[11]。高寒草甸是青藏高原隆升后,长期受高寒气候条件影响所形成的高原地带性植被类型[12],多以寒冷潮湿多年生草本植物群落为主[13]。因不同地形部位、土壤温度、湿度、地下水、土壤性状等的综合影响,使高寒草甸在不同地形部位形成了差异较大的灌丛草甸、嵩草草甸、沼泽化草甸等植被类型,其生产力也表现出明显的差别[14,15]。高寒草甸、高寒灌丛、沼泽化草甸以及山地草原是青藏高原东缘分布面积最广且最为典型的生态系统类型。目前,对青藏高原不同生态系统的植物群落结构[16]、土壤理化性质[6]以及土壤微生物[17]方面的报道较多,但是有关该区域土壤酶活性的研究很少。本研究旨在测定分析不同生态系统土壤中水解酶活性及其与土壤有机碳和全氮含量、含水量等土壤性质的关系,为深入研究青藏高原东缘不同生态系统结构和功能提供科学的基础数据。
1 材料与方法
1.1 研究区概况
研究区域在甘肃省甘南藏族自治州境内,位于青藏高原东部地区(101°~103°77′E,33°36′~35°58′N),海拔2 200~4 000m,属高寒大陆性季风气候;年平均降水量450~780mm,主要集中于7-9月份;年均温1.8℃,1月均温在-10.7℃以下,7月均温11.7℃,生长季最高气温23.6~28.9℃;年平均霜期不少于270d。主要土壤类型为高山草甸土、高山灌丛草甸土、沼泽化草甸土,土壤呈有机质及全氮养分丰富而速效养分贫乏的特点。植物生长期短,生产力较低。本研究中的草地类型主要有高寒草甸(alpine meadow)、高寒灌丛(alpine shrub)、沼泽化草甸(swamp meadow)和山地草原(mountain steppe)。
1.2 土壤样品采集及处理
于2009年8月在甘南州境内用土钻采集土样,具体见表1,所采集的土样分2层(0~15和15~30cm),按照“之”字型取8个土核,除去地上凋落物、石块等杂物后,将8个单样混匀装入密封袋作为1个混合土样(每个草地类型的土样重复数见表1),并立即带回实验室。
每份混合土样分为2部分:第1部分样品风干,其中部分样品过0.15mm筛,用于测定土壤有机碳和全氮含量;另一部分样品过1mm筛,用于测定土壤pH和脲酶活性。第2部分样品采用鲜样,用于测定土壤含水量、酸性磷酸酶活性、β-葡萄糖苷酶活性,过2mm筛后于4℃条件下保存。
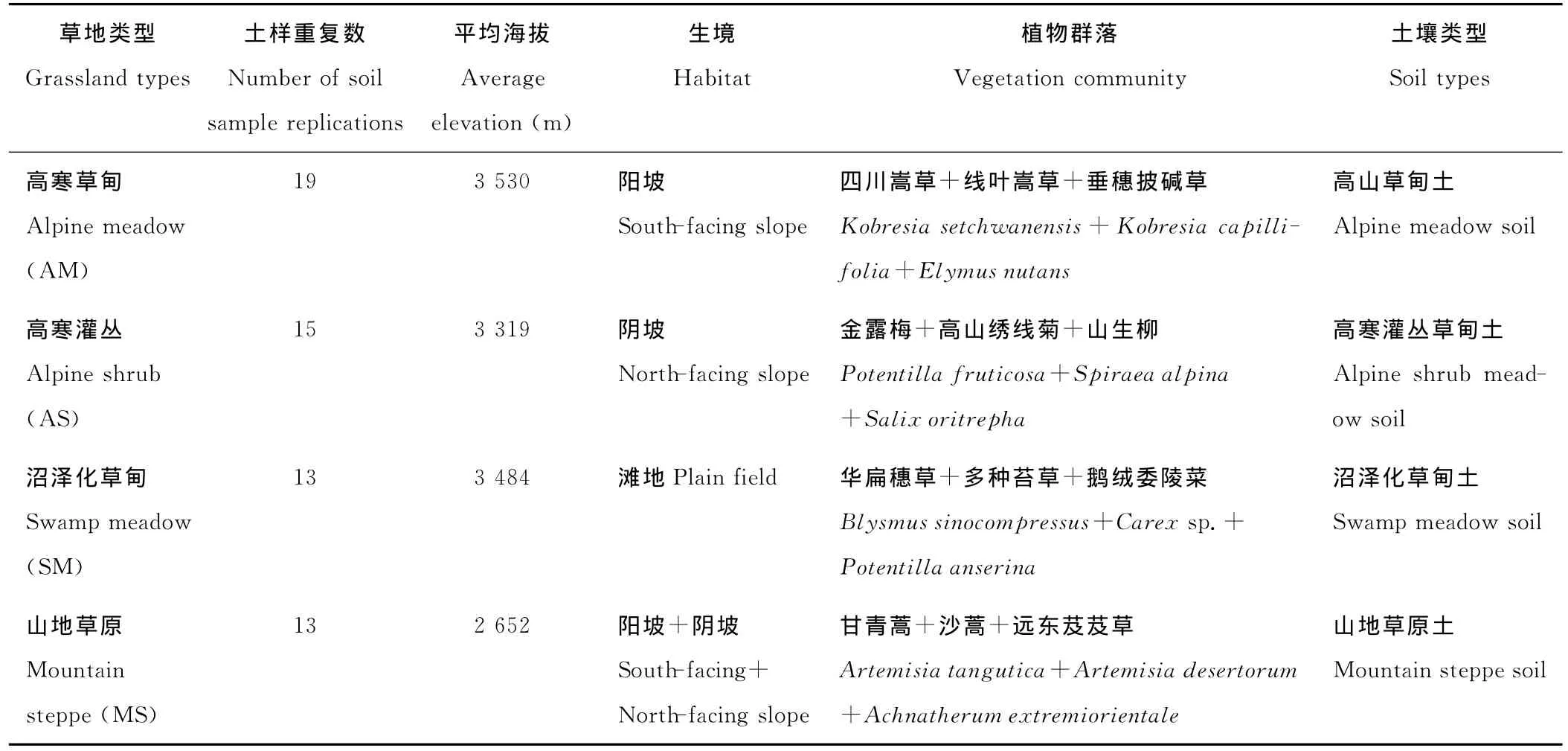
表1 不同草地类型样地基本情况Table 1 General conditions of sample plots of different grassland types
1.3 土壤理化性质的测定
土壤理化性质的测定方法参考文献[18]。土壤有机碳(soil organic carbon,SOC)和全氮(total nitrogen,TN)含量分别用硫酸-重铬酸钾氧化法和半微量凯氏定氮法测定;含水量(%)、pH分别用烘干法、酸度计法[w(土)∶w(去二氧化碳的水)=1.0∶5.0]测定。
1.4 土壤酶活性的测定
土壤脲酶活性采用靛酚蓝比色法[19]测定,其活性以培养24h后每克风干土壤中NH3-N的微克数来表示(μg/g·24h);土壤β-葡萄糖苷酶活性和酸性磷酸酶活性均采用硝基酚比色法[20]测定,其活性以培养1h后每克鲜土中对-硝基酚的毫克数表示(mg/g·h)。
1.5 数据分析
在SPSS 16.0软件中统计分析实验数据。用单因素方差分析检验各种土壤酶活性在相同土层不同草地类型中的差异,用Pearson’s相关分析法分析相关性。
2 结果与分析
2.1 不同草地类型土壤理化性质
各类型草地土壤含水量、有机碳和全氮含量均为表层土高于下层土(表2)。山地草原土壤呈弱碱性,而其他3个草地类型均为弱酸性。含水量和全氮含量在4种类型草地土壤中的变化趋势一致,均为沼泽化草甸>高寒灌丛>高寒草甸>山地草原;有机碳含量的变化趋势为高寒灌丛>高寒草甸>沼泽化草甸>山地草原。
2.2 不同草地类型土壤酶活性
研究表明(图1),土壤β-葡萄糖苷酶、土壤酸性磷酸酶及土壤脲酶活性在不同草地类型间均有显著差异(P<0.05)。除土壤脲酶活性在山地草原基本相同之外,不同草地类型表层土壤(0~15cm)酶活性均高于下层土壤(15~30cm)。土壤β-葡萄糖苷酶活性在高寒沼泽化草甸最高(图1A),0~15和15~30cm土层分别为0.139和0.075mg/(g·h),与其他草地类型差异显著(P<0.05)。不同草地类型表层土壤中的β-葡萄糖苷酶活性依次为沼泽化草甸>高寒灌丛>高寒草甸>山地草原,下层与其相同。土壤酸性磷酸酶活性也在高寒沼泽草甸最高(图1B),0~15和15~30cm土层分别为1.189和0.868mg/(g·h),与其他草地类型差异显著(P<0.05)。不同草地类型表层土壤中的酸性磷酸酶活性依次为沼泽化草甸>高寒草甸>高寒灌丛草甸>山地草原,下层与其相同。土壤脲酶活性以高寒草甸为最低(图1C),0~15和15~30cm土层分别为8.96和7.64μg/(g·24h)。不 同草地类型表层土壤中的脲酶活性依次为沼泽化草甸>高寒灌丛>山地草原>高寒草甸;下层土壤中脲酶活性依次为山地草原>沼泽化草甸>高寒灌丛>高寒草甸。
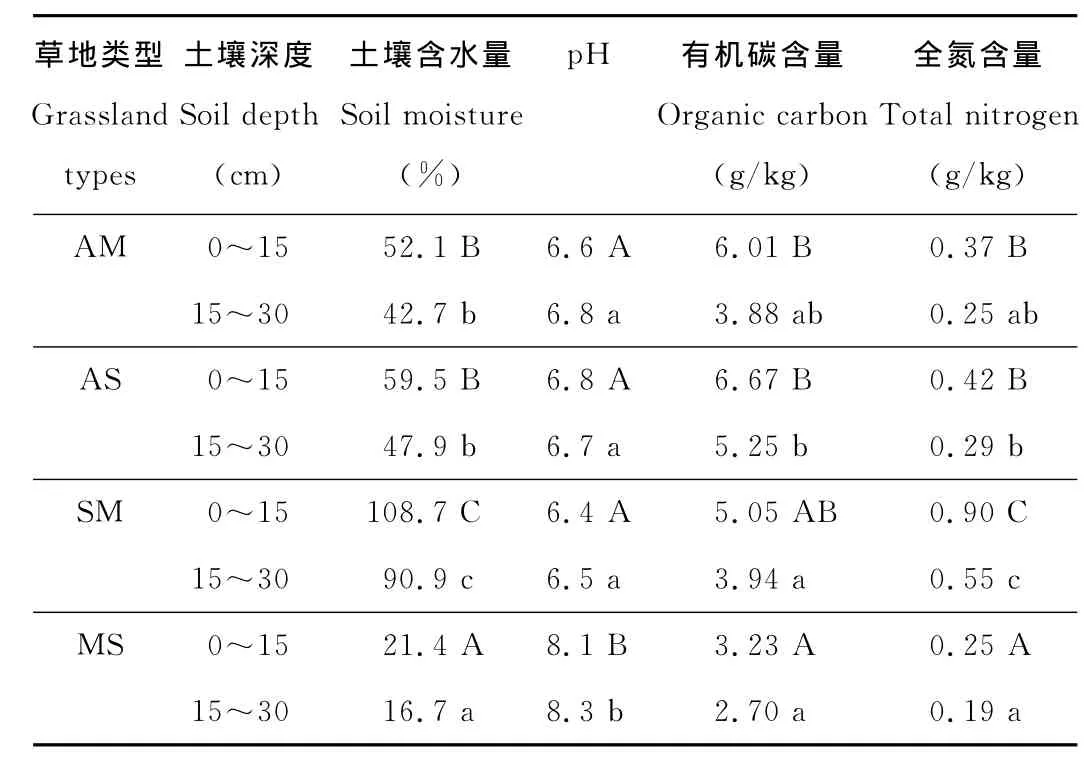
表2 不同草地类型土壤理化性质Table 2 Soil physico-chemical characteristics of different grassland types
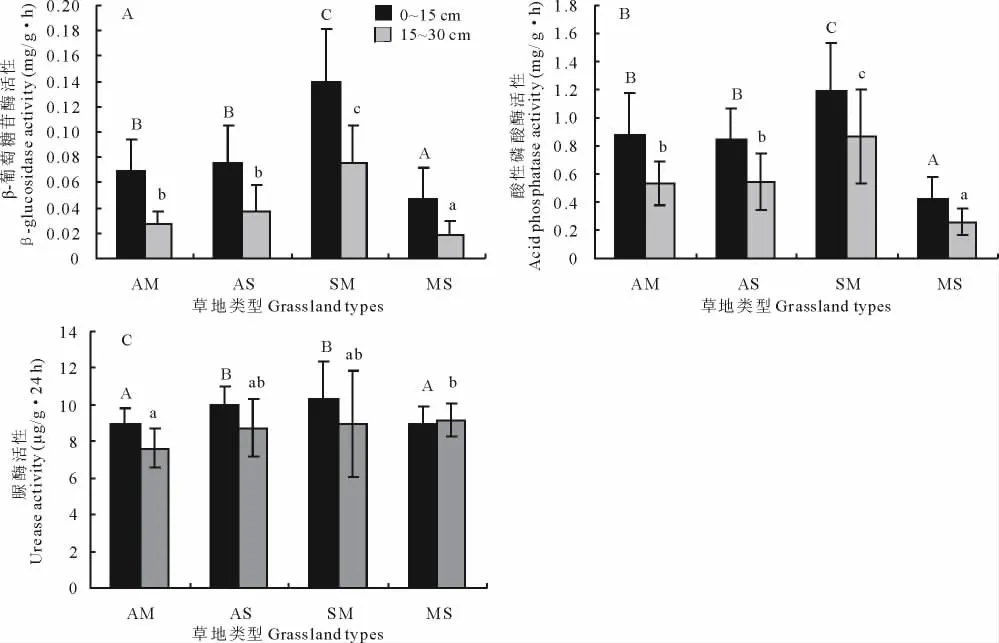
图1 不同草地类型土壤酶活性Fig.1 Soil enzymatic activity of different grassland types
2.3 土壤理化性质与土壤酶活性之间的相关关系
研究结果显示(表3),4种草地类型土壤的β-葡萄糖苷酶活性与有机碳含量和全氮含量呈极显著正相关(P<0.01),与pH呈极显著负相关(P<0.01);土壤酸性磷酸酶活性也与有机碳含量和全氮含量呈极显著正相关(P<0.01),与pH呈极显著负相关;土壤脲酶活性只与全氮含量呈极显著正相关(P<0.01)。
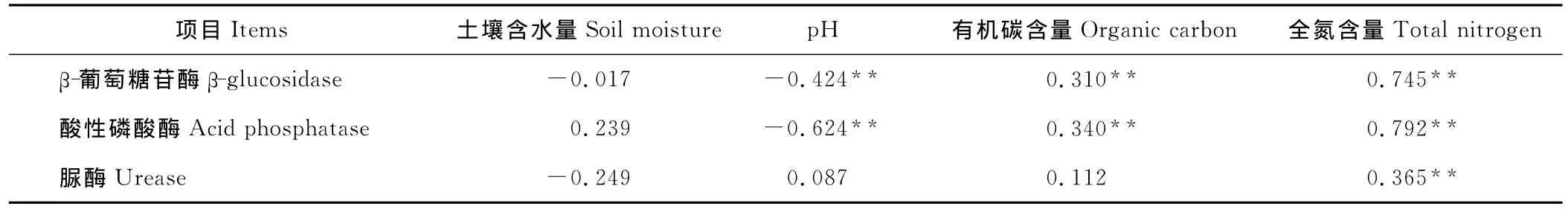
表3 土壤酶活性与土壤理化性质之间的相关分析Table 3 Correlation analysis of soil enzyme activity with soil physical-chemical characteristics
3 讨论
土壤是植物生长的基质,其理化特性决定着植物群落类型的分布[21]。作为对土壤营养异质性分布的反应,植物在养分丰富的局部环境中能选择性地改变其根系的生长,从而增加养分的吸收[22]。4种类型草地中,全氮含量在高寒沼泽化草甸土壤表层和下层都最高,且与其他3种草地类型全氮含量有显著差异,这与王长庭等[6]的结果一致。沼泽化草甸群落地上、地下生物量高于其他植被类型,说明了土壤中养分含量的高低直接影响着群落的生产力,土壤养分越丰富,群落生产力越高[6]。土壤中的有机碳主要取决于土壤中的植物残体量以及土壤微生物作用下分解损失量的平衡状况[23]。4种类型草地中,高寒灌丛土壤表层和下层的有机碳含量都最高,沼泽化草甸土壤中有机碳含量较低,这与杨成德等[24]在东祁连山高寒草地的研究结果一致,但与王长庭等[6]的结果不同。虽然沼泽化草甸中地上地下生物量最丰富,但8,9月份土壤温度和湿度等气候因素适宜时,沼泽化草甸土壤中植物残体分解率比高寒草甸高[25],从而有机碳积累量较低。山地草原由于其植被覆盖度低,土壤水分蒸发较快,土壤有机碳和全氮含量都比其他草地类型中含量低。
土壤酶是土壤中活跃的有机成分之一,在土壤养分循环以及植物生长所需养分的供给过程中起到重要的作用[26,27]。本研究结果表明,土壤酶活性在不同草地类型土壤表层中的含量比下层高,这与多数研究结果一致[6,28]。土壤表层累积了较多的枯枝落叶和腐殖质,有充分的营养源以利于土壤微生物的生长,加之水热条件和通气状况,微生物生长旺盛,代谢活跃,呼吸强度加大而使表层积累了较高的土壤酶活性[28]。土壤β-葡萄糖苷酶是微生物将纤维素分解为葡萄糖的限制酶;土壤磷酸酶将磷酸单酯转化为可被植物吸收利用的无机磷酸盐;土壤脲酶能够酶促土壤中有机化合物尿素分子酰氨碳氮键的水解,生成的氨是植物氮素营养来源之一[19,20]。本研究所测的3种酶都在沼泽化草甸土壤中活性最强,山地草原最弱,原因是沼泽化草甸群落生产力高,为微生物的生长发育提供了较好的碳、氮源[6],从而土壤酶活性也高。土壤β-葡萄糖苷酶和脲酶在高寒灌丛土壤中的活性比高寒草甸高,而酸性磷酸酶活性大小与其相反,说明高寒灌丛土壤中与碳氮元素相关的生化反应比高寒草甸强,而磷素反应则在高寒草甸中较强。山地草原土壤表层与下层脲酶活性基本相同,可能是因其地表多裸露,土壤较干燥,加之植被盖度小,土壤养分贫瘠,土壤从表层到30cm深层之间的生化反应强度变化较小。
土壤中可供植物利用的营养元素的多少,与土壤酶活性的高低直接相关;在良好的有机养分状况下,土壤酶活性较高,其对土壤中营养元素的矿质化作用强度愈大,愈有利于系统内的营养物质循环[27,29,30]。本研究结果表明,脲酶与全氮含量极显著相关,表明其活性可以反映氮素的转化过程[31]。β-葡萄糖苷酶活性与酸性磷酸酶活性均与有机碳含量和全氮含量呈极显著正相关,与pH呈极显著负相关。由此可知,β-葡萄糖苷酶与酸性磷酸酶活性能够反映土壤中有机质和氮素分解状况,可作为衡量土壤肥力的重要指标。
[1]Mersi W,Schinner F.An improved and accurate method for determining the dehydrogenase activity of soils with iodonitrotetrazolium chloride[J].Biology and Fertility of Soils,1991,11:216-220.
[2]Badiane N N Y,Chotte J L,Pate E,etal.Use of soil enzyme activities to monitor soil quality in natural and improved fallows in semiarid tropical regions[J].Applied Soil Ecology,2001,18(3):229-238.
[3]和文祥,朱铭莪.陕西土壤脲酶活性与土壤肥力关系分析[J].土壤学报,1997,34(4):392-398.
[4]杨万勤,钟章成.缙云山森林土壤酶活性的分布特征、季节动态及其与四川大头茶的关系研究[J].西南师范大学学报(自然科学版),1999,24(3):318-324.
[5]杨万勤,王开运.森林土壤酶研究进展[J].林业科学,2004,40(2):152-159.
[6]王长庭,龙瑞军,王根绪,等.高寒草甸群落地表植被特征与土壤理化性状、土壤微生物之间的相关性研究[J].草业学报,2010,19(6):25-34.
[7]Dick R P,Breakwill D,Turco R.Soil enzyme activities and biodiversity measurements as integrating biological indicators[A].In:Doran J W,Jones A J.Handbook of Methods for Assessment of Soil Quality[M].Soil Science Society of America,Madison,1996:242-272.
[8]曾思齐,周国英,佘济云.湘东丘陵区次生林土壤微生物的分布及酶的活性研究[J].中南林学院学报,1998,18(2):20-23.
[9]Decker K L M,Boerner R E J,Morris S J.Scale dependent patterns of soil enzyme activity in a forest landscape[J].Canadian Journal of Forest Research,1999,29(2):232-241.
[10]Edwards C A.Biological Interaction in Soil,in Agriculture,Ecosystem and Environment[M].London:Academic Press,1988:24.
[11]汤懋苍,程国栋.青藏高原近代气候变化及其对环境的影响[M].广州:广东科技出版社,1998.
[12]周兴民,李健华.海北高寒草甸生态系统定位站的主要植被类型及其地理分布规律[A].见:夏武平.高寒草甸生态系统[M].兰州:甘肃省人民出版社,1982:9-18.
[13]王启兰,曹广民,王长庭.高寒草甸不同植被土壤微生物数量及微生物生物量的特征[J].生态学杂志,2007,26(7):1002-1008.
[14]王启基,周兴民.高寒矮嵩草草甸禾草种群的生长发育节律及环境适应性[J].植物生态学与地植物学学报,1991,15(2):168-176.
[15]王启基,王文颖,邓自发.青海海北地区高山嵩草草甸植物群落生物量动态及能量分配[J].植物生态学报,1998,22(3):222-230.
[16]杨时海,李英年,蒲继延,等.三种高寒草甸植被类型植物群落结构及其土壤环境因子研究[J].草地学报,2006,14(1):77-83.
[17]吴建国,艾丽.祁连山3种典型生态系统土壤微生物活性和生物量碳氮含量[J].植物生态学报,2008,32(2):465-476.
[18]鲁如坤.土壤农业化学分析方法[M].北京:农业科技出版社,2000:228-233.
[19]关松荫.土壤酶及其研究法[M].北京:农业出版社,1983:182-266.
[20]吴金水.土壤微生物生物量测定方法及其应用[M].北京:气象出版社,2006:117-137.
[21]王长庭,龙瑞军,刘伟,等.高寒草甸不同群落类型土壤碳分布与物种多样性、生物量关系[J].资源科学,2010,32(10):2022-2029.
[22]Pickett S T A,Kolasa J,Armesto J.The ecological community:A comment on the role of empiricism in ecology[J].Oikos,2002,54:515-525.
[23]齐玉春,董云社,耿元波,等.我国草地生态系统碳循环研究进展[J].地理科学进展,2003,22(4):342-352.
[24]杨成德,陈秀蓉,龙瑞军,等.东祁连山高寒草地牧草返青期土壤酶活性特征[J].草地学报,2010,18(3):308-313.
[25]王启兰,曹广民,姜文波,等.高寒湿地植物残体降解的动态分析[J].草业学报,2004,13(4):39-44.
[26]王光华,金剑,韩晓增,等.不同土地管理方式对黑土土壤微生物量碳和酶活性的影响[J].应用生态学报,2007,18(6):1275-1280.
[27]焦婷,常根柱,周学辉,等.高寒草甸草场不同载畜量下土壤酶与土壤肥力的关系[J].草业学报,2009,18(6):98-104.
[28]文都日乐,李刚,张静妮,等.呼伦贝尔不同草地类型土壤微生物量及土壤酶活性研究[J].草业学报,2010,19(5):94-102.
[29]张焱华,吴敏,何鹏,等.土壤酶活性与土壤肥力关系的研究进展[J].安徽农业科学,2007,35(34):11139-11142.
[30]侯彦会,周学辉,焦婷,等.甘肃永昌县放牧草地土壤脲酶活性与土壤肥力的关系初探[J].草业学报,2009,18(4):111-116.
[31]朱丽,郭继勋,鲁萍,等.松嫩草甸3种主要植物群落土壤脲酶的初步研究[J].植物生态学报,2003,27(5):638-643.
猜你喜欢
成都信息工程大学学报(2022年4期)2022-11-18
青海草业(2022年2期)2022-07-23
生物技术通报(2020年12期)2020-12-21
腐植酸(2020年1期)2020-11-29
活力(2019年21期)2019-04-01
浙江理工大学学报(自然科学版)(2018年6期)2018-11-07
河北林业科技(2016年5期)2016-11-08
现代农业(2016年3期)2016-04-14
土壤与作物(2015年3期)2015-12-08
郑州大学学报(理学版)(2012年4期)2012-03-25
