
间歇性饥饿对瓦氏黄颡鱼生长及肝脏抗氧化功能的影响
2011-09-03 11:30:42安徽农业大学动物科技学院杨严鸥
中国饲料 2011年21期
安徽农业大学动物科技学院 杨严鸥 姚 峰
在鱼类的规模化养殖中,饲料成本的节约是一个重要的问题,一方面可以通过改变饲料配方来节省成本,另一方面,还可以考虑利用补偿生长原理,通过改变投喂策略来节省成本。
补偿生长是指动物受到饥饿、营养不足或其他不良环境条件的胁迫后,在生理上发生应激反应,当环境条件恢复正常、胁迫消失后,这些动物的生长速度会超过未受胁迫的正常个体的现象(Dobson 和 Holmes,1984)。 瓦 氏 黄 颡 鱼 (Pelteobagrus vachelli Richardson)属鲇形目鲿科,是一种杂食偏肉食性的特种经济养殖鱼类,生长速度比黄颡鱼(Pelteobagrus fulvidraco Richardson)快,规格比黄颡鱼大。目前,通过改变投喂方式来促使瓦氏黄颡鱼补偿生长的研究较少,本研究拟对此作一些探讨,旨在为瓦氏黄颡鱼的养殖方式的改变提供参考。
在采用饥饿胁迫产生补偿生长的许多研究中,一般设定的饥饿时间较长(1周以上),连续饥饿后再连续投喂 (Zhu 等,2004;Tian and Qin,2003;Le-Francois等,1999;Paul等,1995)。这些试验着重研究饥饿时间对鱼类补偿生长的影响,或着重研究鱼类在补偿生长中生长率改变的时间历程。本试验采用“饥饿—喂食—再饥饿—再喂食—……”的方式饲养瓦氏黄颡鱼,饥饿时间固定为1 d,投喂时间不等,这种循环饥饿的方式在每次补偿生长减弱时又重新将其激活,有利于不断激发鱼类的补偿生长潜能,增加鱼体补偿生长的时间,对鱼体的伤害也较小 (Mikko等,2009;Wang 等,2009)。
1 材料与方法
1.1 饲养设备 试验在循环水养鱼系统中进行,系统主要包括12只体积为70 cm×55 cm×36 cm的塑料循环水水族箱,每只箱的循环水量为2.00 L/min, 溶氧 >6.2 mg/L,pH值为6.7~7.4,氨氮 <0.09 mg/L。 自然光照,水温(25±1)℃,双制式空调控制。
1.2 试验鱼的驯养及试验饵料 试验鱼取自湖北荆州黄颡鱼良种场,暂养及试验期间均投喂新鲜水蚯蚓。水蚯蚓以微流水暂养,投喂前以聚维酮碘溶液消毒,冲洗干净后再使用。试验期间,每次称量水蚯蚓前用滤纸吸干其水分。同时每天称取部分水蚯蚓作为样本冰冻保存。试验结束后将所有样本混合并烘干,测定其干物质含量为18.50%,干物质中粗蛋白质含量为46.45%,能量为22.36 kJ/g,以代表整个试验期间水蚯蚓的营养水平。
1.3 试验设计 设4种投喂模式:第一种连续投喂水蚯蚓,整个试验期间不间断,为对照组,第二种投喂4 d后饥饿1 d、第三种连续投喂2 d后饥饿1 d、第四种连续投喂1 d后饥饿1 d,4种方式分别以R0、R1/4、R1/2和R1/1表示, 连续投喂是指每天9∶00投过量水蚯蚓,第二天9∶00回收残饵,再投入新鲜的过量水蚯蚓,第三天9∶00再回收残饵,如此循环往复。试验持续45 d(S1/1组循环周期共44 d,在试验开始时多喂1 d,保证试验周期为45 d,与其他组一致),R0、R1/4、R1/2和 R1/1实际投喂天数分别为45、36、30 d和23 d。每一处理含3个平行箱(重复),试验开始前,将鱼饥饿24 h,然后随机取样称重,每箱放入30尾,总初始体重见表1。另取样30尾,分为3份,称重后70℃烘干至恒重,用以分析初始躯体生化成分。
试验结束时,将鱼饥饿24 h,称量每缸鱼总重,再从每缸中随机取10尾鱼分别称重、测体长及内脏重,70℃烘干后测量干重,测定鱼体生化成分。另每缸随机再取10尾鱼,冰浴取肝脏-70℃保存,用于测定抗氧化指标。
试验结束后将残饵称重,并根据溶失率校正整个试验的残饵量。测定溶失率时在3个无鱼的水族箱中投放定量的水蚯蚓,24 h后回收称重,计算溶失率。
1.4 指标测定 测定鱼样及饵料样品的干物质、粗蛋白质、脂肪和能量含量。测定干物质时在105℃下干燥样品至恒重;测粗蛋白质时用元素分析仪(Perkin Elmer 2400,Perkin-Elmer Corporation,Connecticut,U.S.A.)测定含氮量,再乘以 6.25;脂肪采用索氏乙醚抽提法测定;能值采用Phillipson微量能量计(Phillipson microbomb calorimeter,Gentry Instruments Inc.,Aiken,U.S.A.)测定。
SOD用黄嘌呤氧化酶法测定,MDA采用脂质过氧化物测试盒用比色法测定,试剂盒由南京建成生物工程研究所提供,相应操作及计算公式参照说明书进行。
1.5 结果计算
增重率=(Wt-Wo)/Wo;
成活率=100×(总尾数-死亡尾数)/总尾数;
干重特定生长率=100×(lnWdt-lnWdo)/t;
蛋白质特定生长率=100×(lnWPt-lnWPo)/t;
脂肪特定生长率=100×(lnWLt-lnWLo)/t;
能量特定生长率=100×(lnEt-lnEo)/t;
内脏指数=100×内脏重/体重;
肥满度=100×Wt/L3;
平均日摄食量=ITd/t×Wo;
实际日摄食量=ITd/T×Wo;
饵料系数=100×ITd/(Wt-Wo);
平均蛋白质摄食率=200×ITP/t(Wt+Wo);
实际蛋白质摄食率=200×ITP/T(Wt+Wo);
蛋白质储积率=100×(WPt-WPo)/ITP;
蛋白质效率比=(Wt-Wo)/ITP;
平均摄食能=200 ITE/t(Wt+Wo);
实际摄食能=200 ITE/T (Wt+Wo);
能量储积率=100 (Et-Eo)/ITE;
式中 Wt为终末湿重,g;Wo为初始湿重,g;Wdt终末干重,g;Wdo初始干重,g;Lt为体长,cm。 WPt为终末体蛋白重量,g;WPo为初始体蛋白重量,g;WLt为终末体脂肪重量,g;WLo为初始体脂肪重量,g;Et为终末能值,kJ;Eo为初始能值,kJ;t为试验周期,d;T为实际摄食天数,d;ITd为试验期间的摄食量(干重,g),ITP为试验期间的总蛋白质消耗/g,ITE为试验期间的总能量消耗,kJ。
1.6 统计分析 试验数据用平均数±标准差表示。平均数用Duncan’s法进行组间差异的多重比较。
2 结果与分析
2.1 不同投喂模式对生长的影响 由表1知,4种处理之间的成活率、肥满度和内脏指数均无显著差异(P>0.05)。3个试验组的终末体重和增重率均显著高于对照组,出现超补偿生长 (P<0.05),其中R1/4组的超补偿效果最为明显,略高于R1/2组,显著高于R1/1组和对照组。3个试验组的蛋白质、脂肪和能量特定生长率都显著高于对照组 (P<0.05),但3个组间无显著差异 (P>0.05),而4种处理间的干重特定生长率均无显著差异(P > 0.05)。
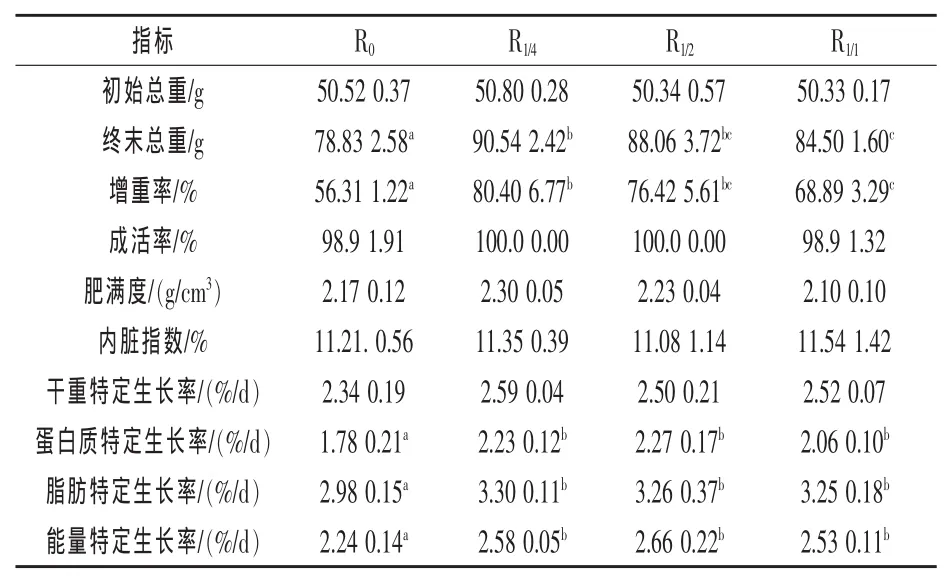
表1 不同投喂模式对瓦氏黄颡鱼的生长、肥满度及内脏指数的影响
2.2 不同投喂模式对饲料利用的影响 由表2知,在整个试验期,饥饿时间越长,平均日摄食量越低,实际日摄食量越高(P<0.05),除处理组R1/4和R1/2实际日采食量差异不显著外,各组间差异均显著(P>0.05)。随着饥饿时间增加,饲料系数显著下降(P < 0.05)。

表2 不同投喂模式对瓦氏黄颡鱼的饲料利用的影响
由表3知,在整个试验期,饥饿时间越长,蛋白质的平均摄食率越低,实际摄食率越高 (P<0.05)。 3个试验组的蛋白质效率比和蛋白质储积率显著高于对照组,其中最高的为R1/1组(P<0.05)。3个试验组中,R1/2组和R1/1组的蛋白质利用效率要显著高于R1/4组(P<0.05),而两者间差异不显著(P>0.05)。饥饿时间越长,平均摄食能越低,实际摄食能越高 (P<0.05)。R1/2组和R1/1组的能量储积率显著高于R1/4组和对照组 (P<0.05),而R1/4组和对照组差异不显著(P>0.05)。
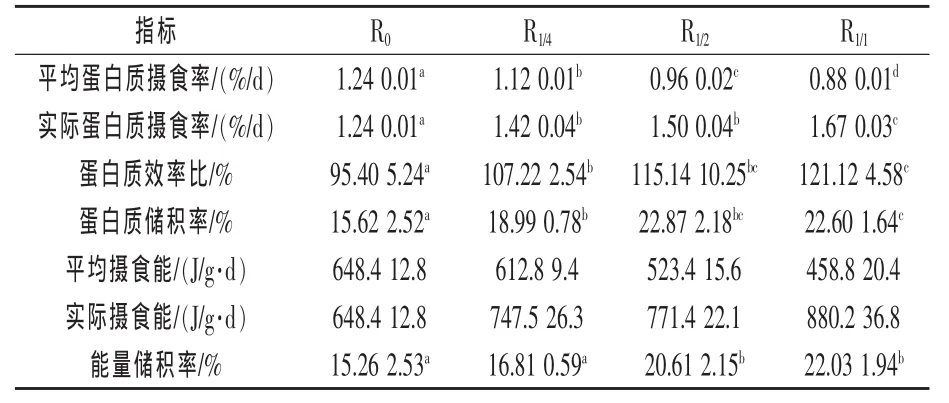
表3 不同投喂模式对瓦氏黄颡鱼的蛋白质和能量利用的影响
2.3 不同投喂模式对抗氧化指标的影响 由表4知,R1/2和R1/1组SOD酶活性显著低于R1/4组和对照组(P<0.05),后两组之间无显著差异;饥饿时间越长,MDA含量越高,其中R1/2组和R1/1组要显著高于R1/4组和对照组(P<0.05)。

表4 不同投喂模式下瓦氏黄颡鱼的SOD酶活性及MD含量
3 讨论
3.1 鱼类在胁迫后的生长按程度可分为超补偿生长、完全补偿生长、部分补偿生长和不能补偿生长。本试验中,R1/4、R1/2和R1/1组出现了超补偿生长现象,其中R1/4组生长效果最好,同时3个试验组的蛋白质、脂肪和能量生长率也显著高于对照组。陈颖(2003)发现,在连续饥饿 5、10、15 d 后再连续投喂,黄颡鱼出现补偿和部分补偿生长现象,没有出现超补偿现象。补偿程度不同,应该与试验设计不同有关,但都表明黄颡鱼是具有补偿生长能力的鱼类。
3.2 鱼类可通过3种方式获得补偿生长:(1)提高饵料转化效率,如美国红鱼(Sciaenops ocellatus)(姜志强等,2002);(2)提高摄食, 如虹鳟(Obcorhynchus mykiss)(Nikki等,2004)和异育银鲫(崔正贺和王岩,2007);(3)同时提高饵料转化效率和摄食,如沟鲶(Ictalurus punctatus)(Kim 和Lovell,1995)、太平洋鲑(Oncorhynchus spp.)(冯健和覃志彪,2006)和千年笛鲷 (Lutjanus sebae)(区又君和刘泽伟,2007)。本试验中,瓦氏黄颡鱼通过同时提高每日的实际摄食量和饵料利用效率(包括蛋白质和能量利用效率)获得了超补偿生长。不过,虽然3个试验组的实际摄食量显著提高,但按整个试验周期(45 d)计算的平均摄食量仍然显著低于对照组,试验组的饲料用量还是比对照组少,饲料得到节约。对饲料利用效率的分析显示,饥饿时间越长,饲料系数越低,蛋白质和能量储积率越高。已有研究显示,补偿强度与鱼体受到的胁迫程度密切相关,在鱼体的承受范围内,补偿强度随饥饿时间的增加而增加 (Jobling等,1994),本试验结果与之一致。
3.3 氧自由基是机体在新陈代谢及杀伤异源物质以及机体抗应激时产生的一类重要物质,机体本身在正常状态下清除过剩的氧自由基的能力是评价机体健康的重要指标。SOD是机体超氧化自由基的清除剂,能阻止并消除自由基的连锁反应,以保护机体免受损害(Demple,1999)。当SOD酶比活力下降时,表明机体的免疫力下降(丁美丽和李光友,1997)。 本试验显示,R1/2组和 R1/1组的SOD酶比活力显著低于对照组和R1/4组,这说明这两种处理中鱼体的机体免疫力受到损害。
丙二醛(MDA)是自由基作用于脂质发生过氧化反应的代谢产物,具有细胞毒性,MDA的高低间接反映了机体细胞受损伤的严重程度(Grundy 和 Storey,1998)。 在胁迫条件下,机体内的丙二醛含量常出现上升现象。例如,温水超过26℃,中华鲟血清的MDA含量显著上升(李大鹏等,2008);饥饿超过一定时间,方斑东风螺肝脏MDA含量显著上升(薛明等,2010)。本试验中,随饥饿时间增加,MDA含量显著上升,其中R1/1和R1/1组显著高于对照组,这反应出试验鱼的机体细胞受到了损害。
4 小结
3个试验组均达到了超补偿生长效果,其中R1/4组增重率最高(P < 0.05),R1/2和 R1/1组的饲料系数虽然低于R1/4组,但平均摄食量较低,因此生长效果差于R1/4组,而且,R1/2和R1/1组的的肝脏抗氧化功能受到损害,因此,综合考虑,采用R1/4的方式可获得好的养殖效果。
[1]陈颖.饥饿和再投喂对黄颗鱼(Pelteobagrus fulvidraco Richardson)生长、耗氧率及生化组成的影响:[硕士学位论文][D].长春:吉林农业大学,2003.
[2]崔正贺,王岩.不同体重异育银鲫 (Carassius auratus gibelio)的补偿生长[J].海洋与湖沼,2007,38(1):8 ~ 14.
[3]丁美丽,李光友.有机污染对中国对虾体内的外环境影响的研究进展[J].海洋与湖沼,1997,28:17 ~ 21.
[4]冯健,覃志彪.淡水养殖太平洋鲑循环饥饿后补偿性生长效果研究[J].水生生物学报,2006,30(5):508 ~ 514.
[5]姜志强,贾泽梅,韩延波.美国红鱼继饥饿后的补偿生长及其机制[J].水产学报,2002,26(1):67 ~ 72.
[6]李大鹏,刘松岩,谢从新,等.水温对中华鲟血清活性氧含量及抗氧化防御系统的影响[J].水生生物学报,2008,32(3):327 ~ 332.
[7]区又君,刘泽伟.千年笛鲷幼鱼的饥饿和补偿生长[J].水产学报,2007,(31)3: 323 ~ 328.
[8]薛明,柯才焕,王德祥,等.饥饿及恢复生长对方斑东风螺抗氧化体系的影响[J].中国水产科学,2010,17(2):281 ~ 288.
[9]Demple B.Radical ideas:genetic responses to oxidative stress[J].Clin Exp Pharm Physiol,1999,26:64 ~ 78.
[10]Dobson S H,Holmes R M.Compensatory growth in the rainbow trout,Saimo gairdneri Richardson[J].J Fish Biol,1984,25:649 ~ 659.
[11]Grundy J E,Storey K B.Antioxidant defenses and lipid peroxidation damage in estivating toads,Scaphiopus couchii[J].J Comp Physiol B,1998,168(2):132 ~ 142.
[13]Jobling M,Meloy O H,Dos S J,et al.The compensatory growth response of the Atlantic cod:effects of nutritional history [J].Aquac Inlernationa ,1994,2:75~ 90.
[14]Le-Francois N R,Blier P U,Adambounou L T,et al.Exposures to lowlevel jonizing radiation;effects on biochemical and whole-body indices of growth in juvenile brook charr[J].J Exp zool,1999,283(3):315 ~ 325.
[15]Mikko K,Juhani P.The effect of intermittent feeding on feed intake and compensatory growth ofwhitesh CoregonuslavaretusL[J].Aquaculture,2009,288:92 ~ 97.
[15]Nikki J,Pirhonen J,Jobling M,et al.Compensatory growth in juvenile rainbow trout,Oncorhynchus mykiss (Walbaum),held individually[J].Aquaculture,2004,235(1 ~ 4):285 ~ 296.
[16]Paul A J,Paul A M,Smith R L.Compensatory growth in Alaska yellowfin sole,Pleuronectes asper,following food deprivation [J].J Fish Biol,1995,46:442 ~ 448.
[17]Russell N R,Wooton R J.Appetite and growth compensatory in European minnows (Phoxinus phoxinuds)following short periods of food restriction[J].Environ Biol Fish,1992,34:277 ~ 285.
[18]Tian X,Qin J.A single phase of food deprivation provoked compensatory growth in barramundi Lates calcarifer[J].Aquaculture,2003,224(1 ~ 4):169~179.
[19]Wang Y,Li C,Qin J G,et al.Cyclical feed deprivation and refeeding fails to enhance compensatory growth in Nile tilapia,Oreochromis niloticus L[J].Aquaculture Research,2009,40(2):204 ~ 210.
[20]Zhu X M,Xie S Q,Zou Z J,et al.Compensatory growth and food consumption in gibel carp,Carassius auratus gibelio,and Chinese longsnout catfish,Leiocassis longirostris,experiencing cycles of feed deprivation and refeeding[J].Aquaculture,2004,241(1 ~ 4):235 ~ 247.
猜你喜欢
当代水产(2021年1期)2021-03-19 05:17:14
防护林科技(2020年9期)2020-11-09 01:45:24
当代水产(2019年9期)2019-10-08 08:02:44
甘肃畜牧兽医(2019年2期)2019-03-20 01:49:42
中南林业调查规划(2018年3期)2018-11-24 03:18:48
文学少年(原创儿童文学)(2016年19期)2016-02-28 17:50:34
文史春秋(2016年8期)2016-02-28 17:41:34
中南林业调查规划(2015年3期)2015-12-20 05:22:31
意林原创版(2015年12期)2015-06-11 01:14:54
应用海洋学学报(2014年1期)2014-11-22 07:17:50
