
饲料中不同比例的鱼粉比对丁鱥幼鱼生长和消化酶的影响
2011-06-29 10:26:52西南大学荣昌校区水产系陈鹏飞何张萍西南大学荣昌校区动物医学系
中国饲料 2011年21期
西南大学荣昌校区水产系 陈鹏飞 何张萍西南大学荣昌校区动物医学系 伍 莉*
蛋白质是水产动物生长的重要营养元素,又是影响饲料成本的重要因子,而作为水产动物饲料中重要的动物蛋白源,由于其资源量下降,价格昂贵,成为影响饲料成本的主要因素之一。因此,寻找适宜的鱼粉比例可以充分利用紧缺的动物蛋白,最大限度地满足养殖对象的生长需求 (陆阳等,2010;江仁党等,2004)。丁鱥(Tinca tinca)属鲤形目鲤科雅罗鱼亚科丁鱥属,是广泛分布于欧洲大陆、亚洲高加索山脉和西伯利亚地区的一种重要的经济鱼类,其含肉率高达73.59%,鱼肉蛋白质含量19.73%,脂肪含量1.29%,氨基酸总量18.13%,其中必需氨基酸量为7.04%(黄峰等,2004)。丁鱥肌肉所含人体必需氨基酸占氨基酸总量的百分比高于WHO/FAO提出标准,必需氨基酸指数为74.38,尤其富含多聚不饱和脂肪酸,二十碳五烯酸含量及二十二碳六烯酸含量在总脂中分别达到了5.14%和6.0%,而被欧洲人视为保健食品(Vacha 和 Eva,1995)。近年来,丁鱥在我国部分地区已有养殖,关于幼鱼生长、营养及饲料的研究已有少量报道(冯新忠等,2008),但有关丁鱥配饲中适宜的蛋白水平还很少报道。本试验以鱼粉作为动物蛋白源,豆柏、菜饼为植物蛋白源来配制试验饲料,旨在探讨丁鱥配饲中适宜的鱼粉比例,为丁鱥应用配合饲料的研制提供参考。
1 材料与方法
1.1 试验鱼 丁鱥幼鱼购于四川隆昌县古宇湖渔场,均重(6.5±0.22)g,共计 900 尾。
1.2 试验饲料 根据丁鱥属肉食性兼杂食性鱼类的特点,饲料以鱼粉作动物蛋白源,豆粕、菜饼为植物蛋白源,并按鱼粉比例占饲料蛋白0%、20%、40%、60%、80%、100%配成 6种饲料,分别投喂试验组丁鱥。为提高饲料的适口性,在饲料中加入少量腥味诱食剂,同时加入适量水产黏合剂,基础日粮配方及营养水平见表1。
1.3 试验设计与饲养管理 试验鱼随机分为6组,每组设3个重复,每个重复50尾,用3%食盐水消毒后养殖于水族箱 (100 cm×48 cm×50 cm)中,暂养1周,待其适应环境、正常摄食后开始试验。24 h充氧,每日投喂2次(09∶00和16∶00),投饲率为3%~5%。试验期间根据养殖水体混浊度采用虹吸法每2~3 d换水一次,每次换水量约占总水量的30%。试验期间水温控制在22~25℃,pH为8.10~8.50,溶氧>7.0 mg/L,氨 <0.30 mg/L,试验时间56 d。试验结束时禁食(自由饮水)24 h后称重,统计每组鱼的生长数据,再与试验前的数据进行比较,即可得出不同比例的鱼粉对试验鱼生长的影响。
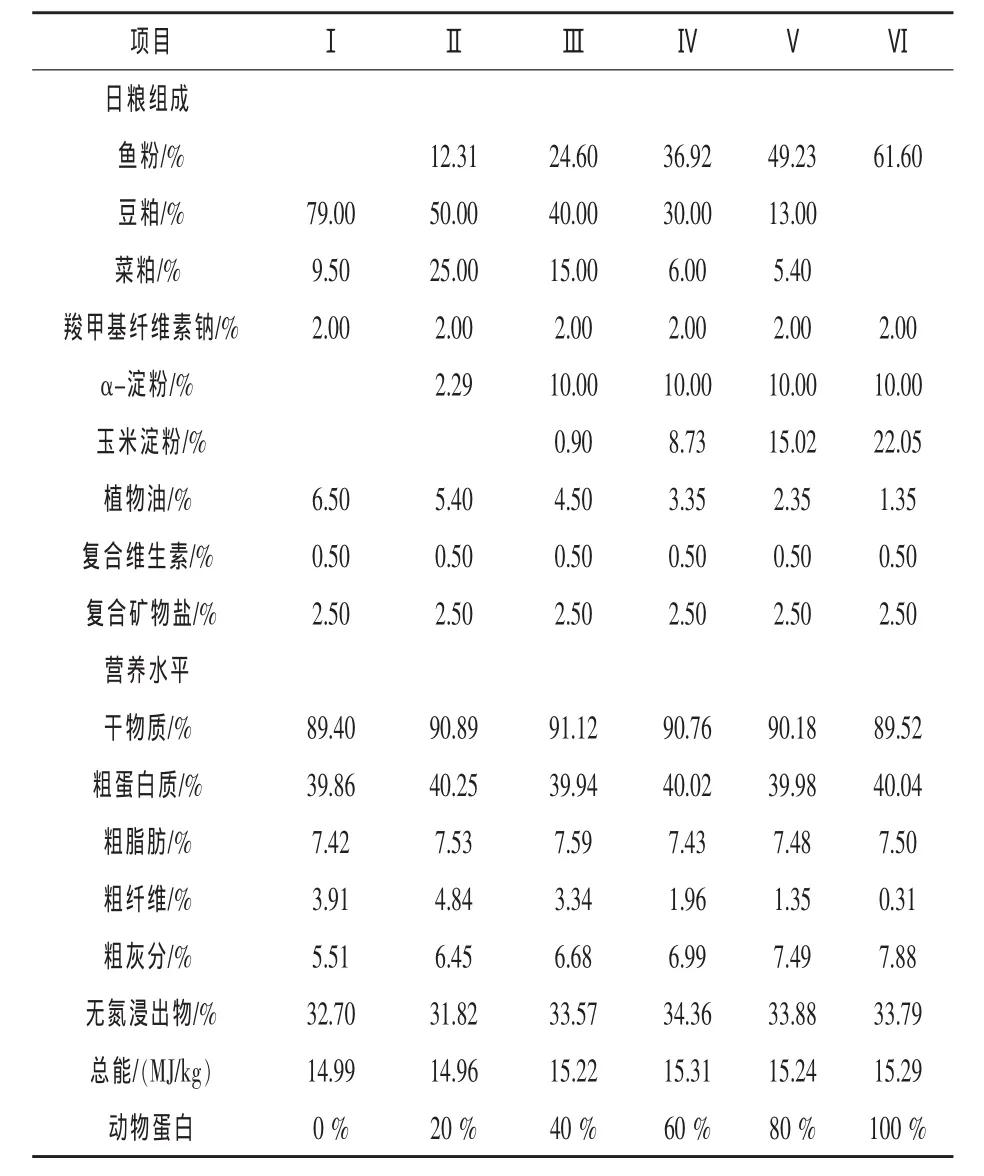
表1 饲料组成及营养水平(理论值)
1.4 生长性能测定 参考李爱杰(1996)和刘永坚等(2002)的方法测定。计算公式如下:
增重率(WGR)/%=100×(终末尾均重-初始尾均重)/初始尾均重;
特定生长率(SGR)/(%/d)=(1n 终末尾均重-ln 初始尾均重)/天数×l00;
饲料系数(FCR)= 摄食量/增重×l00;
蛋白质效率(PER)/%=(终末尾均重-初始尾均重)/蛋白质摄入量×100。
1.5 消化酶活性测定
1.5.1 酶液制备 参照王爱民等(2010)方法制备粗酶提取液,并于24 h内分析完毕。
1.5.2 蛋白酶测定 参照 《生化技术导论》(中山大学生物系,1979)进行。在 pH 7.6,(30±1)℃下保温15 min,以1 min水解酪蛋白产生1 μg酪氨酸为一个活性单位。
1.5.3 淀粉酶 参照 《临床生化检验》(上海市医学化验所,1979)进行。 在 pH 7.6,(30±1)℃下,100 mL酶液在30 min内完全水解10 mg淀粉为一个活性单位。
1.5.4 脂肪酶 参照 《生化技术导论》(中山大学生物系,1979)进行。 在 pH 7.6,(30±1)℃下,脂肪酶水解脂肪1 min产生1 μg分子脂肪酸的酶量定义为一个活性单位。
1.6 数据处理 采用SPSS 17.0对试验数据进行单因素方差分析(One-way ANOVA),差异显著时进行Duncan’s多重比较,P<0.05表示差异显著,P<0.01表示差异极显著。
2 结果
2.1 不同比例的鱼粉对丁鱥幼鱼生长及饲料利用的影响 由表2可见,随着鱼粉比例的提高,丁鱥的增重率、特定生长率和蛋白效率也随之增高,且在鱼粉为60%时达到最大,此时饲料系数为最低。但鱼粉超过60%后,增重率、特定生长率和蛋白效率呈下降趋势,饲料系数开始升高。增重率和特定生长率各试验组之间除第Ⅴ组与第Ⅵ组差异不显著(P>0.05)外,其余各组间差异极显著(P<0.01);饲料系数以第Ⅳ组最低,但与Ⅲ、Ⅴ组比较差异不显著(P>0.05),与Ⅰ、Ⅱ、Ⅵ组比较差异显著(P<0.05);蛋白质效率以第Ⅳ组最高,但与Ⅲ、Ⅴ、Ⅵ组比较差异不显著(P>0.05),与Ⅰ、Ⅱ组比较则差异显著(P<0.05)。
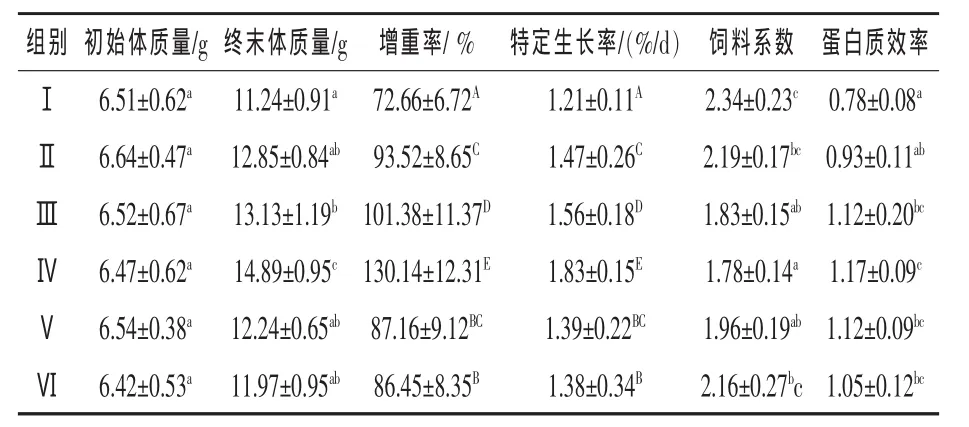
表2 饲料中不同比例的鱼粉对丁鱥幼鱼生长的影响
2.2 不同比例的鱼粉对丁鱥幼鱼消化酶活性的影响 由表3可见,随着鱼粉比例的增加,丁鱥蛋白酶显著升高,并在饲料鱼粉比例为60%时(第Ⅳ组)达到最大,其中肠蛋白酶活性与Ⅰ、Ⅱ、Ⅲ、Ⅵ组比较差异显著(P<0.05),肝胰脏蛋白酶活性差异极显著(P<0.01)。之后,随着鱼粉比例继续增加,蛋白酶活性、淀粉酶活性开始下降。结果显示,饲料中动植物蛋白水平的一定配比可以使肠和肝胰脏蛋白酶活性增加。同时,随着鱼粉比例的增加,丁鱥肠淀粉酶与肝胰脏淀粉酶活性亦呈上升趋势,饲料鱼粉比例在60%时(第Ⅳ组)达到最大(分别为 348.63 U/mg和 443.76 U/mg),但肠淀粉酶活性第Ⅳ组与Ⅲ、Ⅴ组比较差异不显著(P>0.05),与Ⅰ、Ⅱ、Ⅵ组比较差异显著(P < 0.05),第Ⅳ组肝胰脏淀粉酶活性与试验各组比较差异极显著(P<0.01);在相同鱼粉比例条件下,肝胰脏淀粉酶活性高于肠淀粉酶活性,表明适宜的鱼粉比例对激活肝胰脏淀粉酶活性具有促进分泌的作用。鱼粉比例的变化对肠和肝胰脏的脂肪酶活性均无显著影响,各试验组之间差异不显著 (P>0.05)。
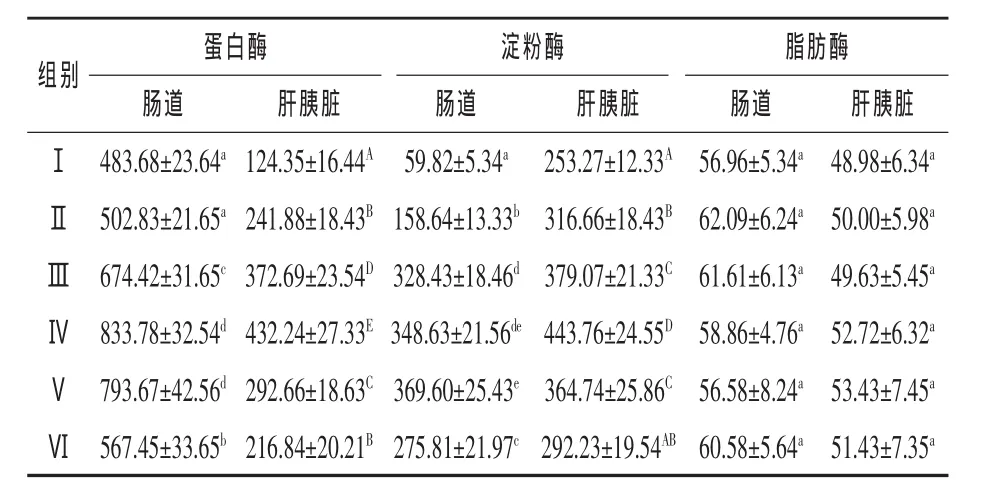
表3 饲料中不同比例的鱼粉对丁鱥肠道及肝胰脏消化酶活性的影响U/mg
3 讨论
3.1 不同比例的鱼粉对丁鱥幼鱼生长及饲料利用的影响 韩庆等(2002)研究发现,随着动物蛋白含量的增高,黄颡鱼增重、蛋白质效率也增高,饲料系数下降,但当动物蛋白含量高于67%时,其增重、蛋白质效率变化不明显,饲料系数反而会略升。中华鲟饲料中动植物蛋白比为1∶1时,幼鲟的生长就能达到较好的效果,当动植物蛋白比达3∶1时,既能发挥中华鲟生长迅速的潜力,又可获得良好的经济效益(庄平等,2002)。但并非动物蛋白含量越高,鱼类生长效果越好 (宋学宏等,2004)。本试验结果表明,随着鱼粉含量从0%上升到60%,试验鱼体增重率、特定生长率也随之增加,达60%时效果最佳,且与其他试验组差异极显著(P<0.01),此时饲料系数最低,蛋白质效率最高,这与动物蛋白具有比植物蛋白更丰富的必需氨基酸等因素密切相关 (周凡和邵庆均,2007)。但当鱼粉含量超过60%后,增重率、特定生长率随之降低,其原因可能有两种:一是由于饲料中鱼粉比例过高,而作为主要植物蛋白源的豆粕比例较少时,虽然能保证饲料总蛋白含量一致,但因饲料中必需氨基酸比例失调而降低了鱼类对饲料的转换率,使其生长速度减缓;二是由于饲料蛋白全由鱼粉提供,或者鱼粉含量过高,在加工和贮存过程中致使鱼粉中的油脂氧化产生毒性,导致鱼类对饲料的利用率降低所致(向枭等,2008)。
3.2 不同比例的鱼粉对丁鱥幼鱼消化酶活性的影响 大量研究表明,饲料蛋白水平在一定范围内,蛋白酶活性随蛋白水平的增加而升高 (石英等,2009;钱曦等,2007;黄峰等,2003)。同时,不同的饲料品质也将影响鱼类的消化酶活性,如鳜鱼胃、肠、肝胰脏蛋白酶活性以饲喂动物性饵料增加幅度最大(孙翰昌,2009)。本试验结果显示,随着饲料中鱼粉含量的提高,能显著提高丁鱥的蛋白酶活性,并在含量为60%时,其蛋白酶活性最高。但随着饲料中鱼粉比例的进一步提高,丁鱥肠和肝胰脏的蛋白酶活性反而呈下降趋势,这与鱼粉含量过高,饲料在加工和贮存过程中致使鱼粉中的油脂氧化,对鱼类产生毒性,导致对消化酶的分泌产生负反馈调节有关(向枭等,2008)。丁鱥肠的蛋白酶活性比肝胰脏的蛋白酶活性高,这与肝胰脏分泌的无活性胰蛋白酶原,在进入肠道后被肠激酶或已有活性的胰蛋白酶所激活有关 (杨蕙萍和童圣英,1998)。
试验结果还显示,随着鱼粉含量从0%~60%增加,肠和肝胰脏的淀粉酶活性逐渐提高。但随着鱼粉比例的进一步增加(超过60%),肠和肝胰脏的淀粉酶活性均有所下降,该结果与其他学者(李芹和刁晓明,2009;王海英等,2008)的研究具有相似性。分析原因,可能是过高的鱼粉用量导致粗纤维含量降低,对肠刺激减弱,从而抑制丁鱥肠和肝胰脏淀粉酶活性。同时,丁鱥肝胰脏的淀粉酶活性比肠高,但两者相差不大,说明丁鱥肠和肝胰脏的淀粉酶对淀粉的消化能力差别不大,在一定程度上证明了鱼类对淀粉的消化是由肠和肝胰脏的淀粉酶共同来完成的(尾崎久雄,1985)。
宋理平等(2010)发现,鱼类的脂肪酶活性与食物组成之间没有明显的相关性。王重刚等(1998)发现,真鲷幼鱼脂肪酶活性与食物中脂肪含量呈负相关趋势。本试验显示,不同比例的鱼粉对丁鱥脂肪酶活性影响并不显著(P>0.05),这与高攀等(2009)和钱曦等(2007)的研究结果一致。究其原因,与饲料中适宜脂肪水平已能满足鱼类对能量的需要所致。
4 结论
综上所述,饲喂不同比例鱼粉的饲料可以影响丁鱥的消化酶活性和生长,当鱼粉含量为60%时,丁鱥肠和肝胰脏蛋白酶和淀粉酶活性最高,增重率、特定生长率、饲料蛋白效率达到最大,饲料系数最低。
[1]冯新忠,苟萍,苏俊,等.额尔齐斯河野生丁鱥营养成分分析[J].食品工业科技,2008,6:276 ~ 279.
[2]高攀,蒋明,文华,等.不同蛋白能量比饲料对草鱼鱼种消化酶活性的影响[J].淡水渔业,2009,39(6):54 ~ 58.
[3]韩庆,罗玉双,夏维福.不同饲料蛋白源对黄颡鱼生长的影响[J].上海水产大学学报,2002,11(3):259 ~ 263.
[4]黄峰,苏德学,田永胜,等.丁鱥的含肉率及其营养价值的分析[J].动物学杂志,2004,39(1):76 ~ 79.
[5]黄峰,严安生,张桂蓉,等.不同蛋白含量饲料对南方鲇胃蛋白酶和淀粉酶活性的影响[J].水生生物学报,2003,27(5):451 ~ 456.
[6]江仁党,林建斌,黄种持,等.不同动植物蛋白比饲料对虹鳟生长的影响[J].淡水渔业,2004,34(3):41 ~ 43.
[7]李爱杰.水产动物营养与饲料学[M].北京:中国农业出版社,1996.89~107.
[8]李芹,刁晓明.不同饵料对瓦氏黄颡鱼稚鱼生长和消化酶活性的影响[J].水生态学杂志,2009,2(1):98 ~ 102.
[9]刘永坚,刘栋辉,田丽霞,等.饲料蛋白质和能量水平对红姑鱼生长和鱼体组成的影响[J].水产学报,2002,26(3):242 ~ 246.
[10]陆阳,杨雨虹,王裕玉,等.不同比例膨化豆粕替代鱼粉对虹鳟生长、体成分及血液学指标的影响[J].动物营养学报,2010,22(1):221 ~ 227.
[11]钱曦,王桂芹,周洪琪.饲料蛋白水平及豆粕替代鱼粉比例对翘嘴红鲌消化酶活性的影响[J].动物营养学报,2007,19(2):182 ~ 187.
[12]上海市医学化验所.临床生化检验(上册)[M].上海:上海科技出版社,1979.366~368.
[13]石英,冷向军,李小勤,等.饲料蛋白水平对血鹦鹉鱼种生长、体组成和肠道蛋白消化酶活性的影响[J].水生生物学报,2009,33(5):874 ~ 880.
[14]宋理平,韩勃,冒树泉,等.脂肪水平对厚唇弱棘鯻生长及肠道脂肪酶活性的影响[J].广东海洋大学学报,2010,30(1):13 ~ 17.
[15]宋学宏,凌去非,蔡春芳,等.丁鱥的营养素需要量及饲料最适能量蛋白比[J].饲料工业,2004,25(9):53 ~ 56.
[16]孙翰昌.不同饲料对鳜生长性能及消化酶活性的影响 [J].中国饲料,2009,24:12 ~ 35.
[17]王爱民,吕富,杨文平,等.饲料脂肪水平对异育银鲫生长性能、体脂沉积、肌肉成分及消化酶活性的影响[J].动物营养学报,2010,22(3):625 ~633.
[18]王海英,孙谧,薛长湖,等.大菱鲆配合饲料中植物蛋白替代鱼粉的可行性研究[J].海洋科学,2008,32(6):9 ~ 12,34.
[19]王重刚,陈品健,顾勇,等.不同饵料对真鲷稚鱼消化酶活性的影响[J].海洋学报,1998,20(1):103 ~ 106.
[20]吴尚忠译(尾崎久雄著).鱼类消化生理(下册)[M].上海:上海科学技术出版社,1985.390~398.
[21]向枭,周兴华,陈建,等.动植物蛋白比对鲤鱼消化酶活性的影响[J].粮食与饲料工业,2008,7:37 ~ 39.
[22]杨蕙萍,童圣英.国内外关于水产动物消化酶研究的概况[J].大连水产学院学报,1998,9(3):64 ~ 69.
[23]中山大学生物系.生化技术导论[M].北京:人民教育出版社,1979.52~54.
[24]周凡,邵庆均.不同蛋白源替代鱼粉的研究进展[J].饲料研究,2007,6:9~11.
[25]庄平,陈喜斌,曾翠平,等.中华鲟幼鲟饲料中适宜动植物蛋白比的研究[J].动物营养学报,2002,14(1):61 ~ 64.
[26]Vacha F,Eva T.Content of polyunsaturated fatty-acids and cholesterol in muscle tissue of tench (Tinca tinca),common carp(Cyprinus carpio)and hybrid of bighead carp (Aristichthys nobilis)with silver carp (Hypophthalmichthys molitrix)[J].Polskie archiwum hydrobiology,1995,42(1 ~ 2):152 ~ 157.
猜你喜欢
当代水产(2022年8期)2022-09-20 06:46:06
当代水产(2022年6期)2022-06-29 01:13:12
当代水产(2021年9期)2021-12-02 01:35:10
当代水产(2021年8期)2021-11-04 08:49:02
医学前沿(2021年18期)2021-04-14 02:53:06
意林·全彩Color(2019年9期)2019-10-17 02:26:00
当代水产(2019年6期)2019-07-25 07:52:08
当代水产(2019年1期)2019-05-16 02:41:50
科学与财富(2018年8期)2018-05-09 01:30:10
广东饲料(2016年2期)2016-12-01 03:43:06
