
宝天曼国家级自然保护区栎类天然林林分空间结构特征1)
2011-08-09 11:07:50闫东锋吴明作杨喜田
东北林业大学学报 2011年9期
闫东锋 吴明作 杨喜田
(河南农业大学,郑州,450002)
维持天然林森林生态系统处于健康状态是天然林保护的首要目标,是发挥其在生物多样性保护和生态功能的前提。我国现有大部分天然林均遭受不同程度的干扰,保持原始状态的森林已经十分少见,恢复其原有结构,调整和改造现有林分结构,人工促进其演替为健康稳定的群落,是天然林经营中亟待解决的问题。对林分结构方面的研究,有许多报道[1-2],这些研究着重于以植物个体数量为基础的树种结构、径级结构和垂直结构等方面,而林分空间结构能够反映林分内树种的空间关系[3-4],决定着林分内光照和温度等小环境的分布,对林木生长和森林经营方式的选择等都有着重要影响,因此,林分的空间结构研究在现代森林经营中具有十分重要的意义。同时,在对保护区进行保护和开发过程中,必须重视制定合理的森林经营措施。宝天曼国家级自然保护区处于北亚热带向暖温带过渡区,虽然前人[5]对其群落结构进行了大量研究,但有关林分空间结构的研究未见报道。笔者利用混交度、角尺度、大小比数及由参照树与4株最近相邻木构成的结构单元的平均距离,研究了处于不同发育阶段的栎类天然林空间结构变化特征及其差异,为保护和经营该地区天然林资源提供借鉴。
1 研究区概况
宝天曼国家级自然保护区位于河南省西南部,秦岭东段,伏牛山南坡。地理位置为北纬 33°25'~33°33',东经 111°53'~112°,总面积为53.4 km2。土壤以山地棕壤为主,土层厚度40~100 cm。该区共有高等植物223科、1 002属、2 711种及变种,分别占河南省高等植物科的87%、属的74%、种的57%。主要乔木树种有锐齿栎(Quercus acutidentata)、短柄枹树(Quercus glandulifera var.breripetiolata)、栓皮栎(Quercus variabilis)、白桦(Betula platyphylla)、化香(Platycarya strobilacea)、三桠乌药(Lauraceae obtusiloba)和漆树(Toxicodendron vernicifluum)等。
2 研究方法
2.1 样地调查与数据处理
在调查区选择林相完整、有代表性的14块样地,面积为20 m×20 m,用Topcon全站仪进行每木定位,同时记录样方内胸径大于4.5 cm的所有乔木个体的种名、胸径、树高、枝下高和冠幅等。对每块样地,将全站仪测定的每木定位坐标图导入到地理信息系统软件Mapgis7.0中。采用点抽样方法,从一个随机点开始,间隔一定距离,设置一个抽样点,记录距离抽样点最近的4株胸径大于4.5 cm个体(参照树)的种名和距离。间隔距离确定的原则是,抽样点4株相邻木不应重复。同时调查参照树与相邻木构成的结构单元的树种数。由于很难获得每株树真实可靠的年龄,本文参考其他学者的方法和经验[6-7],用大小径级结构代替龄级结构:4.5 cm≤d<9.5 cm为Ⅰ龄级;9.5 cm≤d<14.5 cm为Ⅱ龄级;d≥14.5 cm 为Ⅲ龄级。
2.2 内业分析
混交度(M)为参照树的n株最近相邻木中与参照树不属同种的个体所占的比例:

式中:当参照树i与第j株相邻木为非同种时,Vij=1,否则Vij=0。Mi=0,零度混交;Mi=0.25,弱度混交;Mi=0.5,中度混交;Mi=0.75,强度混交;Mi=1,完全混交。
计算混交度平均值时,采用修正的混交度[8-10]计算公式:

式中:N为调查的林木株数;n'i为第i株树所处的结构单元中的树种个数。
角尺度[9](W)是描述相邻木围绕参照树均匀性的指标,为α角小于标准角α0的个数占所考察的4个夹角的比例,有:

式中:当第j个α角小于标准角时,Zij=1,反之,Zij=0;当Wi=0、0.25、0.5、0.75、1 时,分别表示绝对均匀分布、均匀分布、随机分布、不均匀分布和团状分布。
大小比数(U)为胸径大于参照树的相邻木数目占全部最近相邻木的比例:

式中:若相邻木j比参照树i大时,kij=1,否则,kij=0。
空间结构参数在龄级间的差异显著性检验采用K—S检验方法[10],计算公式为:

式中:^D为2个待检验样本之间的最大绝对差异值;^F1与 ^F2分别为2套样本各自的累计频数;n1与n2分别为2套样本各自的样本数。
临界值 Dα=0.05为:
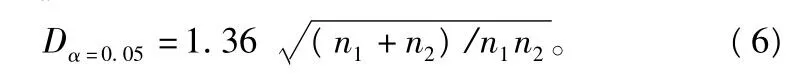
当 ^D > Dα=0.05,差异显著;否则,差异不显著。
3 结果与分析
3.1 混交度
根据式(1)和式(2),统计各样地所有参照树的混交度,以及各龄级和整个林分平均混交度的频率分布,结果见表1。从表1可知:龄级越高,零度混交和弱度混交的林分所占比例越小,Ⅰ龄级和Ⅱ龄级平均混交度峰值出现在弱度混交下,分别为0.318和0.311;Ⅲ龄级在中度混交分布频率最大,为0.320。总体来看,龄级越大,混交程度越大。在该地区,低龄级的林分以栎类为主,占有绝对优势,严重抑制了其他树种的生长发育。而随着林分的发育,生态位相近的栎类种间相互竞争,发生自疏现象,使最近邻体中同种个体数量迅速减少,其他树种逐步占据并取代由于同种竞争而被淘汰的林木个体位置。
3个龄级在强度混交和完全混交的林分所占比例均不高,且基本相同。这说明林分年龄越小,纯林所占的比例越高,林分年龄越大,越容易出现中度混交现象,但3个龄级强度混交林分所占比重均较低,平均混交程度不高。对各龄级平均混交度的K—S检验结果表明,差异不显著(D=0.064,Dα=0.05=0.108)。这是因为幼龄林以栎类萌生林为主,密度较大,不利于多物种共生,只有通过适度间伐才能培育混交度较高的林分。

表1 各龄级混交度的频率分布
3.2 角尺度
表2显示的是各龄级角尺度频率分布,¯W为各龄级所有参照树的平均角尺度。Ⅰ龄级平均角尺度值为1的频率较Ⅱ龄级、Ⅲ龄级明显偏高,林木间的竞争较为激烈;各龄级平均角尺度在0和1上出现的频率均较少,而在0.5的尺度上,分别占到各自结构单元总数的51.1%、55.4%、44.8%,即处于绝对均匀和团状分布这两种极端状况的结构单元很少出现,说明参照树周围最近相邻木出现随机分布的情况最多。当地林分为天然林皆伐后天然更新的林分,人为干扰较少,保护相对完整,可认为是自然演替的结果,林木主要呈随机分布现象。
K—S差异显著性检验结果表明,3个龄级的林分平均混交度差异不显著(D=0.054,Dα=0.05=0.108)。根据角尺度判定标准[8]:平均角尺度在0.475~0.517时为随机分布,小于0.475时为均匀分布,大于0.517时为聚集分布。由表2知:Ⅰ、Ⅱ、Ⅲ龄级的林木平均角尺度均在0.475~0.517范围内,林分整体呈随机分布状态;Ⅰ龄级平均角尺度较Ⅱ、Ⅲ龄级大,位于随机分布的下限临界值附近,聚集程度比后两者偏高。

表2 各龄级角尺度的频率分布
3.3 大小比数
表3反映了不同龄级主要树种胸径平均大小比数以及各龄级所有参照树胸径平均大小比数,3个龄级中,锐齿栎平均大小比数分别为 0.331、0.469、0.549。在以锐齿栎为参照树构成的结构单元中,锐齿栎处于优势地位,但随着龄级增加,锐齿栎的优势地位有所下降。在Ⅰ龄级Ⅱ龄级中,除锐齿栎、短柄枹树、栓皮栎外,其他树种的平均大小比数都在0.5及其以上,而栓皮栎只在Ⅲ龄级中出现,且平均大小比数为0.563,说明除锐齿栎处于绝对优势地位外,其他树种在结构单元中受相邻树种抑制。同时,随着龄级增加,锐齿栎、三桠乌药、短柄枹树的平均大小比数呈递增趋势,其中,三桠乌药在Ⅰ龄级的平均大小比数比较高,即在胸径生长上较其它林木处于劣势,且随龄级增加,劣势地位进一步加剧。

表3 各龄级胸径大小比数的频率分布
3.4 空间结构单元距离尺度
为了进一步分析宝天曼国家级自然保护区栎类天然林空间结构单元中林木的分布状态,对6个在当地分布较多的树种,按龄级分别计算出其参照树与4株最近相邻木的平均距离及标准偏差,计算结果如表4。随着龄级增加,林分平均距离呈现递增的趋势,从1.90 m增加到2.57 m,主要树种亦表现同样的趋势。如:从Ⅰ龄级到Ⅲ龄级,锐齿栎平均距离从1.86 m 增加到 2.11 m,化香从 1.80 m 增加到 2.44 m,这意味着林分及主要树种通过自然稀疏,林木间竞争逐渐减弱,但幅度较小。

表4 各龄级主要树种结构单元的平均距离 m
从表4可以看出,随着龄级增加,各参照树与最近相邻木的平均距离逐渐加大;Ⅰ、Ⅱ龄级结构单元平均距离均分布在[1 m,3 m)区间内;Ⅲ龄级结构单元平均距离均分布在[2 m,3 m)距离组内,且平均距离小于1 m的结构单元没有出现。因此,根据各龄级林分生长发育特点,对距离较近的弱势林木需要进行抚育间伐,在不同距离组内保留相应的林木数,人工促进合理的空间结构状态的形成。
4 结论与讨论
通过对宝天曼国家级自然保护区正在恢复过程中的栎类天然林空间结构对比分析发现:林分总体上处于弱度混交水平,低龄级的混交度水平没有高龄级的高,且锐齿栎在林分中占有绝对优势地位,其他树种均处于受压状态,但随着龄级增加,锐齿栎的优势地位下降;林分总体呈现随机分布状态,低龄级林分聚集程度比高龄级的高,随着林木生长发育,空间结构单元的平均距离呈递增趋势,但平均距离处于3 m以内的结构单元占绝对优势,林木之间的竞争仍十分剧烈。
宝天曼国家级自然保护区是我国中部地区植物群落类型最复杂、植物种类最丰富的地区,天然次生林分是其主要的林分类型,在进行森林经营过程中,应以调整到合理的林分空间结构为目的,通过调整现有林分的树种组成、水平分布格局和林内树种竞争关系,改变树种单一的现状,降低锐齿栎的优势地位,改善其他伴随树种的生长环境。本研究表明,宝天曼国家级自然保护区低龄林分的聚集程度较高,林木空间结构单元平均距离较低,这不利于林分的生长,应通过抚育间伐减少林木之间的竞争,促进林分的正常发育,加速演替的进行,促进形成随机分布的林分以及较高的群落生物多样性,从而使组成林分的树木个体和整个林分群体均朝健康方向发育。
[1]邓送求,闫家锋,关庆伟,等.紫楠风景林林分空间结构分析[J].东北林业大学学报,2010,38(4):29-32.
[2]惠刚盈.角尺度——一个描述林木个体分布格局的结构参数[J].林业科学,1999,35(1):37-42.
[3]龚直文,亢新刚,顾丽,等.长白山云冷杉针阔混交林演替过程空间格局变化[J].东北林业大学学报,2010,38(1):44-56,53.
[4]夏富才,赵秀海,潘春芳,等.长白山阔叶红松林林分空间结构[J].应用与环境生物学报,2010,16(4):529-534.
[5]闫东锋,何静,郭芳,等.宝天曼栎类天然次生林乔木层种间联结性研究[J].西北林学院学报,2009,24(1):26-30.
[6]徐海,惠刚盈,胡艳波,等.天然红松阔叶林不同径阶林木的空间分布特征分析[J].林业科学研究,2006,19(6):687-691.
[7]田国恒,王德强,谷建才,等.塞罕坝机械林场不同林分类型空间结构的比较[J].北京林业大学学报,2010,32(3):80-83.
[8]赵中华,惠刚盈,袁士云,等.小陇山锐齿栎天然林空间结构特征[J].林业科学,2009,45(3):1-6.
[9]闫家锋,邓送求,周伟,等.空间结构单元的侧柏林群落特征分析[J].东北林业大学学报,2010,38(3):1-3,7.
[10]王威,郑小贤,梁雨,等.北京八达岭林场黄栌风景林空间结构分析[J].东北林业大学学报,2008,36(2):25-26,39.
猜你喜欢
湖南生态科学学报(2022年4期)2022-11-11 03:44:34
河南农业大学学报(2020年2期)2020-05-22 02:56:30
山西林业科技(2019年1期)2019-05-27 12:24:52
三明学院学报(2016年2期)2016-10-14 08:43:29
石油化工(2015年9期)2015-08-15 00:43:05
东北林业大学学报(2014年8期)2014-08-02 03:54:12
现代电子技术(2014年13期)2014-07-09 18:34:01
无机化学学报(2014年9期)2014-02-28 17:33:05
中国石油大学学报(自然科学版)(2014年2期)2014-02-28 16:12:24
